Pain Transmission
The sensation, delivery, and interpretation of pain sensation are complex. The discussion below will highlight the mechanisms involved, which have clinical implications of how patients sense/describe pain as well as provide a basis for pharmacologic treatment.
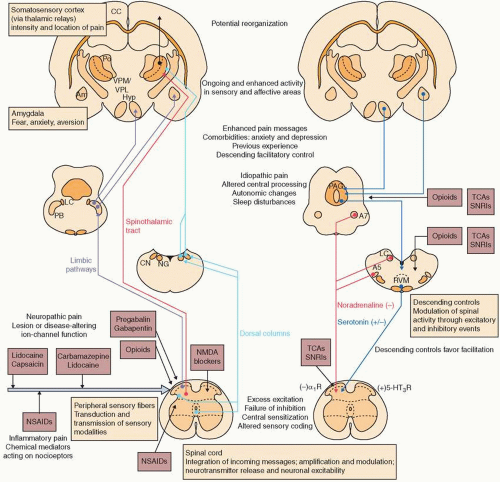
In its most simplified form, nociception signals begin in primary afferent neurons, which are subsequently transmitted through the dorsal horn of the spinal cord and along ascending pathways to the thalamus and somatosensory cortex. Primary afferents are the principal sensory neurons for nociception. The cell bodies of these sensory neurons are located in the dorsal root ganglia (DRG) or in the corresponding ganglion of specific cranial nerves (
Fig. 11-1).
10 As sensory neurons can be very diverse, they are categorized in the Erlanger-Gasser classification according to the size of the axon and presence or absence of myelin. The three major categories are, in order from largest diameter to
smallest, A, B, and C (
Table 11-1).
10 Neurons of Groups A and B are coated with myelin, which enhances nerve conduction speed. Group A is further subdivided into four subgroups, of which Aβ and Aδ are important to pain sensation and signaling. Aβ provide cutaneous touch and pressure afferents, whereas Aδ are mechanoreceptors, nociceptors, thermoreceptors, and sympathetic postganglionic fibers.
10 Group B are sympathetic preganglionic fibers.
10 Group C are small, unmyelinated mechanoreceptors, nociceptors, thermoreceptors, and sympathetic postganglionic fibers.
10The primary afferents that contribute to pain sensation are Aδ and C fibers, and each transmits different types of pain sensation. As Aδ are myelinated, they transmit acute pain rapidly, typically sensed as “sharp” by a patient. C fibers are small and unmyelinated, and thereby transmit pain in a more delayed fashion, often described as “burning” in
quality. Free nerve endings of both Aδ and C fibers respond to varied stimuli intensity including tissue injury, possible tissue injury, and even threatened injury.
10 This graded response may be intended to prevent injury, although it may also contribute a mechanism to development of chronic pain states.
Cell bodies for both Aδ and C fibers are located in the DRG of the spinal cord or respective cranial nerves they serve. Most of the sensory neurons terminate in the ipsilateral dorsal horn. However, the spinal neurons bifurcate into ascending and descending branches to innervate several spinal segments. The dorsal horn is divided anatomically into locations known as Rexed laminae. Aδ and C fibers converge on lamina I (marginal zone), lamina II (substantia gelatinosa), and lamina X (central canal). Lamina V comprises wide dynamic range neurons. In lamina V, Aβ, Aδ, and C fibers converge, which leads to the phenomenon of “referred pain.” An example of referred pain is how visceral pain from myocardial infarction can be felt as pain in the left arm.
In the dorsal horn, the sensory nerves connect with the second-order neurons of the spinothalamic and spinoreticulothalamic tracts. Sensory nerves to the spinothalamic tract cross the contralateral ventrolateral tract and ascend the ventral horn to the thalamus. Sensory nerves to the spinoreticulothalamic tract ascend ipsilaterally in the ventrolateral tract, have medullary projections, and finally ascend to the thalamus.
From the thalamus, a nociceptive signal is transmitted to the somatosensory cortex for discriminative sensation, or to the anterior cingulate gyrus or inferior insula for sensation of the affective-motivational sensation of pain. These pathways are responsible for the affective phenomenon of how pain is experienced physically and emotionally.
Anatomically, the vascular supply of the spinal cord has important clinical implications. Although the posterior spinal cord is supplied by two posterior spinal arteries, the anterior spinal cord is supplied by a solitary anterior spinal artery. The largest anterior spinal artery, the artery of Adamkiewicz, has variable anatomic locations and is vulnerable to injury or spasm. Infarction of the anterior spinal cord is a devastating complication that leads to paraplegia with intact proprioception.
Mechanisms of Neuropathic Pain
Understanding the mechanisms of pain is important, as the sensation described by the patient can identify the type of injury/aberrancy that governs treatment. Mechanisms involved in neuropathic pain are multiple and overlapping. As such, a single patient may report multiple mechanisms of pain with one nidus or injury. This understanding supports the clinical importance of multimodal analgesia for treatment of pain.
As the description of pain is important to understanding its mechanism, the International Society of Pain developed a taxonomy of pain descriptors (
Table 11-2; http://www.iasp-pain.org/Taxonomy). These descriptors can be divided into positive and negative symptoms, based on the response of the patient to stimuli. Positive painful symptoms include allodynia (pain generated by a nonpainful stimulus, such as light touch), hyperalgesia (increased pain from a painful stimulus), and hyperpathia (raised threshold or heightened pain response). Positive symptoms that may or may not be associated with pain include dysesthesia (unpleasant abnormal sensation, spontaneous, or evoked), paresthesia (abnormal sensation, may not be unpleasant, spontaneous, or evoked), and hyperesthesia (increased sensitivity to stimulation, but may not be painful). Negative symptoms include hypoalgesia (decreased sensitivity to painful stimulus) and hypoesthesia (decreased sensitivity to stimulus, not necessarily painful stimulus).
There are at least five accepted mechanisms for neuropathic pain that explain the sensations described above. These mechanisms include ectopic activity, peripheral sensitization, central sensitization, impaired inhibitory modulation, and activation of microglia (
Table 11-3).
11 Ectopic activity is thought to be secondary to hyperexcitability
and spontaneous firing of neurons occurring after an insult, thereby explaining spontaneous symptoms such as paresthesia and dysesthesia; such neuropathic pain can be constant, intermittent, or paroxysmal. Ectopic activity has been attributed to changes in voltage-gated sodium and potassium channels, and hyperpolarization of cyclic nucleotide-gated channels.
11Peripheral sensitization results from hyperexcitability and reduced activation threshold of primary afferent neurons, described by the patient as hyperalgesia or allodynia.
11 Peripheral sensitization may be thought of as inflammatory pain, and may be due to changes in the transient receptor potential cation channel subfamily V member 1 (TRPV1) ion channel.
12Central sensitization is classified as central neuroplastic changes involving spinal or supraspinal nerves. The patient may describe exaggerated response to painful stimulus, hyperalgesia, allodynia, and/or anticipation. Aβ touch fibers develop phenotypic changes including increased neuropeptide expression of substance P and increased excitatory amino acid transmission via
N-methyl-D-aspartate (NMDA) receptors.
11 These changes may explain some of the purported analgesic benefit of NMDA modulating agents such as ketamine (see below). Microglial activation occurs following injury by upregulation of chemokine receptors and release of glial cytokines and growth factors.
11 These changes are believed to contribute to central sensitization and increased pain sensation reported by the patient.
Inhibitory neurons project from the central nervous system (CNS) and serve to modify painful stimuli. Apoptosis of spinal inhibitory interneurons has been reported as a consequence of some injuries, which may explain abnormalities in inhibitory neuron signaling.
13 Without modulation from inhibitory neurons, a peripheral painful stimulus may be interpreted by the CNS as pain of significantly increased severity.