Abstract
Basement membranes represent highly specialized matrices at the interface of cells and stroma. As adhesive units composed of intricately organized intracellular, transmembrane, and extracellular proteins, basement membranes provide tissues with mechanical support and compartmentalization. They also act as a substrate for cell attachment and migration, facilitate tissue repair, and influence the morphogenesis, differentiation, and apoptosis of epithelial cell layers. This chapter’s focus is the epidermal basement membrane, including the structure and function of hemidesmosome-anchoring filament complexes, the lamina densa, and the sublamina densa region. An overview is also provided of bullous diseases that result from acquired or inherited alterations in structural proteins within the epidermal basement membrane.
Keywords
epidermal basement membrane, hemidesmosome, lamina lucida, anchoring filaments, lamina densa, anchoring fibrils, sublamina densa, integrin, laminin, collagen, epidermolysis bullosa, immunobullous diseases, salt-split skin
▪ Bullous pemphigoid antigen 1: BP230 ▪ Bullous pemphigoid antigen 2: BP180; type XVII collagen ▪ Laminin 311: laminin 6 ▪ Laminin 332: laminin 5; epiligrin; kalinin; nicein; GB3 antigen; BM600 ▪ Laminin 511: laminin 10 ▪ Type VII collagen: the epidermolysis bullosa acquisita antigen ▪ Nidogen: entactin
- ▪
Basement membranes serve as: (1) a substrate for attachment of cells; (2) a template for tissue repair; (3) a matrix for cell migration; (4) a substratum to influence differentiation, morphogenesis, and apoptosis of epithelial cell layers; and (5) a permeability barrier for cells and macromolecules
- ▪
By transmission electron microscopy, the major ultrastructural subregions (from superior to inferior) of the epidermal basement membrane are: (1) the cytoskeleton, hemidesmosomal plaques, and plasma membranes of basal keratinocytes; (2) an electron-lucent region termed the lamina lucida; (3) the lamina densa; and (4) the sublamina densa region of the papillary dermis
- ▪
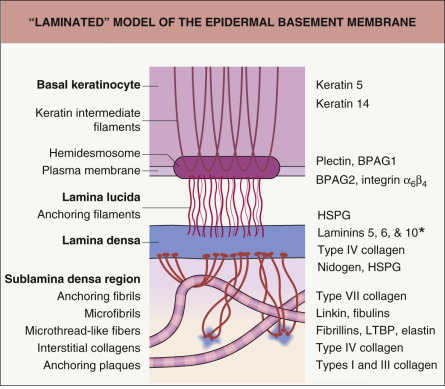
In the “laminated” model of the epidermal basement membrane, keratin intermediate filaments within basal keratinocytes attach to small (i.e. <0.5 microns), electron-dense units (hemidesmosomes) on the basal plasma membranes of these cells. In turn, anchoring filaments (small thread-like strands) connect hemidesmosomes to the lamina densa. The lamina densa is tethered to the dermis by anchoring fibrils, units that by originating and ending in the underside of the lamina densa create ultrastructural loops that serve as attachment sites for fibrillar proteins in the papillary dermis
- ▪
Acquired or inherited abnormalities in structural proteins within the epidermal basement membrane often result in a disease phenotype characterized by blister formation
Introduction
Basement membranes are specialized structures located between cells and their underlying stroma or between different cell types . Basement membranes of various tissues differ ultrastructurally, biochemically, and functionally. All basement membranes contain an electron-dense, somewhat granular matrix of variable thickness that is referred to as the lamina densa. Major components of the lamina densa include type IV collagen, laminins, and heparan sulfate proteoglycans. Other components of the lamina densa vary depending upon the tissue type. Basement membranes serve a variety of functions that are manifest in a tissue-specific manner. Roles common to virtually all basement membranes include acting as: (1) substrates for attachment of cells; (2) templates for tissue repair; (3) matrices for cell migration; (4) substrata to influence differentiation, morphogenesis, and apoptosis of epithelial cell layers; and (5) permeability barriers for cells and macromolecules. The two major basement membrane complexes in skin are found at the epidermal–dermal interface and around the dermal microvasculature. This chapter will deal largely with the epidermal basement membrane.
The epidermal basement membrane is a highly specialized structure that contains numerous tissue-specific elements ( Table 28.1 ). The first characterization of this ultrastructural region in the skin was made via electron microscopy, which identified four distinct subregions of the epidermal basement membrane ( Fig. 28.1 ). These subregions, from superior to inferior distribution, are:
- •
the cytoskeleton, hemidesmosomal plaques, and plasma membranes of basal keratinocytes
- •
an electron-lucent region called the lamina lucida that contains delicate filaments connecting hemidesmosomes (HDs) in basal keratinocytes to the underlying lamina densa
- •
the lamina densa
- •
the sublamina densa region containing anchoring fibrils, anchoring plaques, and filamentous proteins of the papillary dermis.
| REPRESENTATIVE ADHESION PROTEINS IN THE EPIDERMAL BASEMENT MEMBRANE |
| Cytoskeleton of basal keratinocytes |
|
| Hemidesmosome-anchoring filament complexes |
|
| Lamina densa |
|
| Sublamina densa region |
|
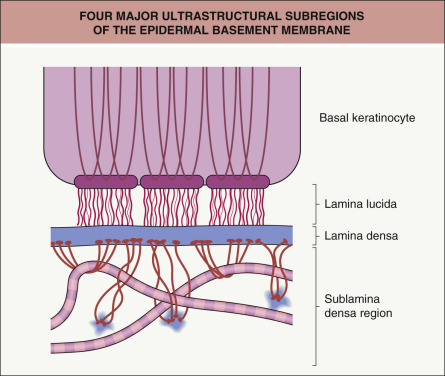
These subregions of the epidermal basement membrane, readily identified in fixed skin samples by transmission electron microscopy, have served as a useful conceptual model of a highly complex adhesion unit that preserves the integrity of the skin ( Fig. 28.2 ). Interestingly, specialized transmission electron microscopy studies of cryopreserved skin have suggested that the lamina lucida may actually represent an artifact of tissue dehydration . Nonetheless, the classic laminated model of the epidermal basement membrane serves as a useful framework to guide understanding of its structure, function, and impairment in disease.
Origin of the Epidermal Basement Membrane
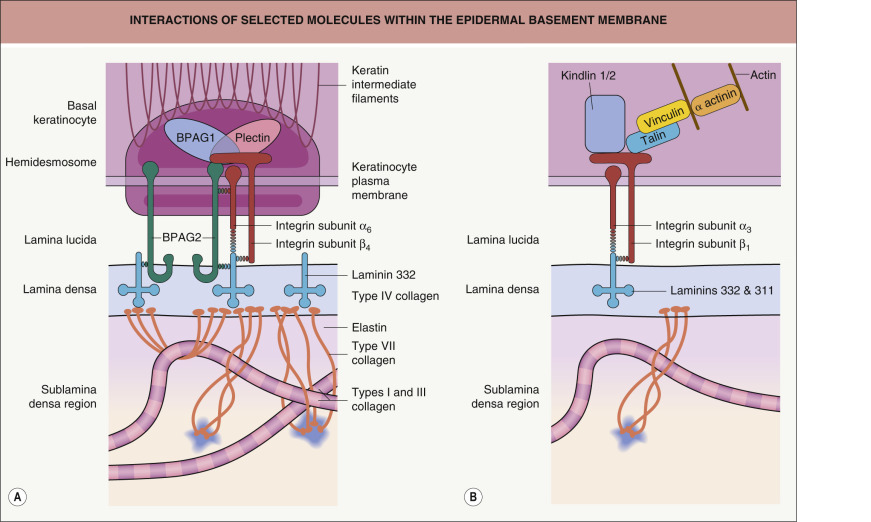
The epidermal basement membrane is comprised of proteins derived from keratinocytes of ectodermal origin as well as from dermal fibroblasts of mesodermal origin. Basal keratinocytes produce hemidesmosomal components such as plectin, the epidermal isoform of bullous pemphigoid antigen 1 (BPAG1e), bullous pemphigoid antigen 2 (BPAG2; collagen XVII), and integrin subunits α 6 and β 4 ; they also contribute CD151 tetraspan, types IV and VII collagen, laminins 332 (α3β3γ2) and 311 (α3β1γ1), and heparan sulfate proteoglycans to the epidermal basement membrane. Dermal fibroblasts produce nidogen, types IV and VII collagen, and other proteins that are translocated to the plasma membranes of basal keratinocytes where they condense and are incorporated within the basement membrane. Integrins and other proteins within the plasma membrane of basal keratinocytes are thought to provide key localization and organizational cues that facilitate the addition of these fibroblast-derived proteins to the basement membrane. The major proteins that comprise the various subregions of the epidermal basement membrane are described below ( Fig. 28.3 ).
Basal Keratinocytes: Hemidesmosome-Anchoring Filaments and Other Integrin Complexes
Plectin
Plectin is a 500 kDa, dumbbell-shaped, dimeric protein associated with the cytoplasmic plaque of HDs . Like BPAG1, desmoplakin, periplakin, and envoplakin (see Ch. 29 ), plectin is a member of the plakin family. Plakins are large, multidomain cytolinkers that connect cytoskeletal networks to each other and to adhesive junctions such as desmosomes and HDs .
The carboxy terminus of plectin binds keratin and vimentin intermediate filaments, whereas its amino terminus contains domains that bind the cytoplasmic tail of integrin subunit β 4 , BPAG2, and actin. Mutations in the gene encoding plectin are responsible for epidermolysis bullosa (EB) simplex associated with muscular dystrophy as well as for the Ogna type of EB simplex and a rare subtype of EB simplex with pyloric atresia (see Ch. 32 ). Targeted inactivation of the gene encoding plectin impairs the cytoarchitecture of skin, muscle, and heart in mice .
Bullous Pemphigoid Antigen 1
Circulating autoantibodies from patients with bullous pemphigoid (BP) were used to define two autoantigens that reside in HDs of basal keratinocytes. The first of these autoantigens characterized, BPAG1e, is a 230 kDa plakin protein localized to the cytoplasmic plaque of HDs . It has a central coiled-coil α-helical rod domain that contains a regular periodicity of acidic and basic amino acid residues. Studies showing that the periodicities of these residues are 180 degrees out of phase suggested that these proteins may self-aggregate by ionic interactions. The globular carboxy terminus of BPAG1e contains acidic and basic amino acids with a periodicity that promotes association with keratin intermediate filaments. The amino terminus of BPAG1e associates with the cytoplasmic domain of BPAG2, integrin subunit β 4 , and ERBIN. The ERBIN protein interacts with the transmembrane tyrosine kinase receptor Erb-B2, thus serving as a potential link between HD biology and Erb-B2 signaling .
Consistent with the role that BPAG1e plays in cytoskeletal organization, BPAG1 knockout mice show signs of epithelial fragility in basal keratinocytes . Unexpectedly, these mice also demonstrate neurologic impairment characterized by dystonia and ataxia, which results from concomitant inactivation of neuronal isoforms of BPAG1 referred to as BPAG1n or dystonin. The amino termini of neuronal isoforms of BPAG1 differ from their epidermal counterpart in that BPAG1n contains actin- or microtubule-binding domains essential for maintaining the cytoarchitecture of neurons. Homozygous nonsense BPAG1 mutations that affect only the epithelial isoform underlie a form of EB simplex .
Bullous Pemphigoid Antigen 2
BPAG2 (collagen XVII) is a type II transmembrane collagen associated with HD-anchoring filament complexes in basal keratinocytes . The cytoplasmic amino terminus of BPAG2 consists of approximately 500 amino acid residues, containing several potential phosphorylation sites within its central portion. The extracellular domain of BPAG2 contains 15 interrupted repeats that have multiple glycine-X-Y amino acid sequences; the latter are characteristic of proteins in the collagen family and therefore are referred to as collagenous domains (coll 1–15; see Fig. 31.9 ). By rotary shadowing, the intracytoplasmic domain of BPAG2 appears as a globular head and the extracellular region as a central rod (corresponding to coll 15) with a flexible tail (corresponding to coll 1–14) . Immunoelectron microscopy studies suggest that the coll 15 rod domain of BPAG2 inserts into the lamina densa and that its carboxy-terminal tail then loops back through the lamina densa into the lamina lucida (see Fig. 28.3 ) .
BPAG2 occurs in two forms : (1) a 180 kDa full-length protein; and (2) a 120 kDa extracellular domain that is shed from plasma membranes of basal keratinocytes by proteolysis. Proteolytic processing of BPAG2 is mediated by “sheddases” of the disintegrin and metalloproteinase (ADAM) family, in particular the tumor necrosis factor-α-converting enzyme (TACE). Phosphorylation of BPAG2 by ecto-casein kinase 2 inhibits its proteolysis by TACE, a mechanism whereby the adhesion and motility of adjacent basal keratinocytes may be modulated .
In the epidermal basement membrane, the extracellular domain of BPAG2 is found as a homotrimer in which the collagenous domains form a cross-linked triple helix. The helix forms independently of amino acid sequences in the amino terminal portion of BPAG2 and is apparently organized in a 5′ to 3′ orientation using the 16th n on- c ollagenous portion of the protein (NC16) as a nucleation site . The cytoplasmic domain of BPAG2 associates with BPAG1e, integrin subunit β 4 , and plectin . The first extracellular segment of BPAG2 (NC16) contains 73 amino acid residues that are thought to bind integrin subunit α 6 . Immunogold electron microscopy studies have suggested that the carboxy-terminal portion of BPAG2 interacts with laminin 332 at the interface of these two proteins near the junction of the lamina lucida and the lamina densa (see Fig. 28.3 ) .
BPAG2 is targeted by autoantibodies from patients with BP, pemphigoid gestationis (PG), mucous membrane pemphigoid (MMP), and linear IgA bullous dermatosis (LABD) . Autoantibodies from patients with BP, PG, and LABD typically target the NC16A domain of BPAG2 (see Fig. 31.9 ), whereas those from patients with MMP tend to target the distal carboxy terminus of BPAG2, which extends deeper into the basement membrane, as well as NC16A. Patients with less severe subtypes of junctional EB often possess null mutations in the gene encoding BPAG2 ( COL17A1 ) ( Table 28.2 ). These individuals characteristically demonstrate a complete lack of BPAG2 in their epidermal basement membrane, along with a tendency for skin fragility, subepidermal blister formation, alopecia, dystrophic nails, and dental enamel hypoplasia . Interestingly, COL17A1 mutations resulting in deletion of the intracellular domain of BPAG2 that associates with BPAG1, plectin, and integrin subunit β 4 result in a phenotype characterized by intraepidermal as well as junctional blister formation .
| TARGETS COMMON TO AUTOIMMUNE AND INHERITED BLISTERING DISEASES | |||
|---|---|---|---|
| Protein target | Structural target | Autoimmune disease | Genetic disease |
| BPAG1e | HD | BP | Recessive EB simplex |
| Type XVII collagen (BPAG2) | HD-anchoring filament complexes | BP, PG, MMP, linear IgA bullous dermatosis | Junctional EB (often milder) |
| Integrin subunit β 4 | HD-anchoring filament complexes | Ocular MMP | Junctional EB with pyloric atresia |
| Laminin 332 | Lamina lucida–lamina densa interface | Anti-epiligrin MMP | Junctional EB (often more severe) |
| Type VII collagen | Anchoring fibrils | EB acquisita Bullous eruption of SLE | Dystrophic EB (dominant and recessive) |
Integrins
Integrins are heterodimeric transmembrane receptors that promote cell–cell and cell–matrix interactions . Ligand binding to integrin receptors modulates cell adhesion, signal transduction, gene expression, growth, and other fundamental biologic processes. All integrins are αβ-heterodimers. Numerous α- and β-subunits have been identified; several of these have alternatively spliced cytoplasmic domains (e.g. α 1 , α 3 , α 4 , α 6 , α 11b ). Although some of the α-subunits can associate with more than one β-subunit, there is restriction in the number of heterodimers that actually form.
Both α- and β-integrin subunits are glycoproteins with a single hydrophobic transmembrane domain and a cytoplasmic domain that is typically composed of ≤50 amino acid residues, although integrin subunit β 4 has a larger cytoplasmic domain of >1000 amino acid residues. Cytoplasmic domains of integrins typically interact with the actin cytoskeleton (see below) and other intracellular proteins. The cytoplasmic domains and associations of integrin α 6 β 4 represent an exception to this model in that they are concentrated within HDs of epithelial cells .
The extracellular domains of integrin subunits associate to form dimers; subunits do not rely upon associations in their cytoplasmic or transmembrane regions for dimerization. Both subunits of the heterodimer are required for ligand binding. In fact, α- and β-subunits are thought to form a pocket that serves as a specific ligand-binding site. Switching either the α- or β-subunit can change ligand specificity. Individual integrin heterodimers can often bind to more than one ligand, and, similarly, individual ligands are often recognized by more than one integrin. Integrins usually bind specific sequences of peptides in a given group of adhesion molecules, such as the Arg-Gly-Asp (RGD) sequence present in fibronectin and vitronectin. Integrin ligand binding is divalent cation-dependent, and the cation itself (e.g. Ca 2+ , Mg 2+ , Mn 2+ ) can affect the affinity and the ligand specificity of the integrin.
Integrin-mediated cell–cell and cell–matrix adhesion triggers clustering of integrins and the assembly of signaling and adapter proteins at their cytoplasmic domains. This creates an “outside in” signaling complex that conveys biochemical messages to different cellular compartments and links the integrin adhesion site to the actin cytoskeleton. In addition, an “inside out” signaling process fine-tunes affinity for extracellular ligands by altering the conformation of integrin ectodomains. Proteins that serve as major “inside out” integrin activators include talins and kindlins . In basal keratinocytes, kindlin-1 and -2 (also known as fermitin homologs 1 and 2) and other proteins such as vinculin can form integrin-tethered macromolecular complexes termed focal adhesions through which the actin cytoskeleton connects to the extracellular matrix (see Fig. 28.3B ).
Mutations in the FERMT1 gene encoding kindlin-1 are responsible for Kindler syndrome, an autosomal recessive disease characterized by trauma-induced blistering, photosensitivity, poikiloderma, variable mucosal involvement, and increased risk of mucocutaneous squamous cell carcinoma (see Ch. 32 ) . Depletion of kindlin-1 in human keratinocytes results in defective signaling by Rho GTPases involved in remodeling of the actin cytoskeleton and impaired adhesion due to reduced activity of the β 1 integrin subunit. It was recently found that loss of kindlin-1 in mouse keratinocytes also leads to decreased α v β 6 integrin-mediated transforming growth factor-β (TGF-β) activation and increased Wnt signaling, thereby stimulating cutaneous stem cell proliferation and potentially contributing to skin cancer risk as well as cutaneous atrophy due to stem cell exhaustion .
Hemidesmosome-associated integrin α 6 β 4
The large cytoplasmic tail of integrin subunit β 4 contains sequences that appear to be required for HD assembly. The membrane-proximal region of integrin subunit β 4 directly associates with plectin, while its distal carboxy-terminal region binds BPAG2 (see Fig. 28.3 ) . Integrin subunit α 6 exists in two forms, A and B; the former predominates in keratinocytes. The proximal extracellular domain of integrin subunit α 6 binds the NC16A region of BPAG2 . Using K562 cells stably expressing α 6A β 4 and α 6B β 4 , investigators demonstrated that laminins 111 and 332 serve as the preferred ligands for this HD-integrin .
Patients harboring biallelic mutations in genes encoding either α 6 or β 4 integrin subunits develop junctional EB associated with pyloric atresia. Such individuals typically have subepithelial blistering of the oral and respiratory epithelia as well as the skin. Targeted deletions of genes encoding murine integrin subunits α 6 or β 4 also result in extensive subepithelial blisters of the same tissues. Evidence of nuclear and cytoplasmic degeneration in basal keratinocytes of β 4 -null mice suggests that this integrin subunit plays a role in cell survival . Transgenic mice carrying a targeted deletion of the cytoplasmic domain of integrin subunit β 4 have a proliferative defect of stratified and simple epithelia . However, targeted deletion of integrin subunit α 6 does not result in obvious developmental problems in murine epithelia.
Plasma membrane-associated integrins
Non-hemidesmosomal integrins expressed in basal keratinocytes include integrins α 2 β 1 and α 3 β 1 . The former is located predominantly along lateral and apical aspects of basal keratinocytes; the latter is found in the same distribution as well as along the basal aspect of these cells’ plasma membranes. Integrins bind the actin cytoskeleton and are thought to play a key role in cell–cell adhesion. Keratinocytes in culture utilize integrin α 3 β 1 to mediate initial adhesion to extracellular matrix via interaction with laminin 332.
Though integrin α 3 β 1 does not play a role in HD assembly, it is thought to provide key signals regulating cytoskeletal assembly, function of integrin α 6 β 1 , and the establishment and/or maintenance of basement membrane integrity . Mice with targeted deletion of integrin subunit α 3 have disorganized basement membranes in the kidney and skin . While the lamina densa in these mice is missing (or sparse) at sites between HDs, that beneath their HDs appears to be normal. Mild blistering in mice lacking integrin subunit α 3 further substantiates the idea that this polypeptide plays an important role in maintaining the integrity of the epidermal basement membrane and assembly of extracellular matrix. Recently, homozygous mutations in the gene encoding the α 3 integrin subunit were found to underlie a human disorder characterized by basement membrane disruption resulting in congenital nephrotic syndrome, interstitial lung disease, and a mild form of EB .
Tetraspan CD151
CD151 is a cell surface protein that belongs to the tetraspan superfamily. Tetraspan proteins, named for their four membrane-spanning domains, form “web-like” networks with other membrane proteins and have roles in regulating a variety of cellular processes. To date, CD151 is the only tetraspan known to be associated with HDs. In human skin, CD151 associates with integrins α 6 β 4 in HDs and α 3 β 1 in the basolateral plasma membrane of basal keratinocytes. Immunoelectron microscopy studies show that CD151 is concentrated in HDs.
In integrin subunit β 4 -deficient keratinocytes, CD151 clusters with α 3 β 1 at the basal cell surface. Introduction of integrin subunit β 4 into the latter cells yields α 6 β 4 integrins that become incorporated into α 3 β 1 –CD151 clusters, where they are thought to induce the formation of HDs. Soon thereafter, the amount of α 3 β 1 integrins in such clusters diminishes, and CD151 becomes predominantly associated with α 6 β 4 integrins within HDs. CD151 is therefore considered to be a component of pre-HDs whose recruitment is regulated by integrin α 6 β 4 . CD151 is thought to provide a framework for the spatial organization of different HD components .
The Lamina Densa
Laminins
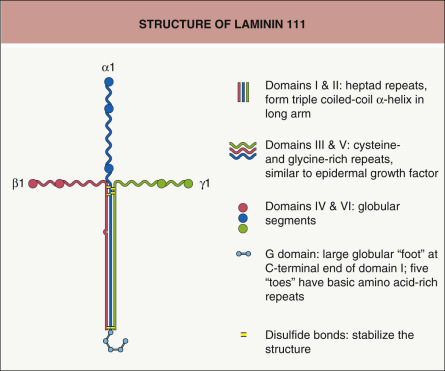
Laminins are a family of heterotrimeric glycoproteins composed of at least 15 members . Laminins consist of three subunits (α, β, and γ) that are joined and stabilized by interchain disulfide bonds and other associations ( Fig. 28.4 ). Each laminin subunit is encoded by a different gene. To date, five α-, three β-, and three γ-subunits have been characterized . Of note, the laminin α3 subunit gene ( LAMA3 ) encodes two transcripts, α3A and α3B, corresponding to short and long variants, respectively.
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


