Abstract
Human papillomavirus (HPV) infections of skin and mucosa are very common and in general cause benign papillomas or warts. More than 150 HPV types have been completely sequenced, and more types probably exist. Most infections are transient, subclinical, and cleared by a cellular immune response. Persistent infection with sexually transmitted high-risk mucosal types, mostly HPV-16 and -18, cause all cervical and most anal cancers, a subset of vaginal, vulvar, penile and oropharyngeal cancers, and rarely cancers of the digits. The three licensed subunit vaccines (bivalent, quadrivalent, and 9-valent) are comprised of empty virus-like particles (VLP); when administered prophylactically, they are highly efficacious in preventing type-specific persistent HPV infection and neoplasia. Epidermodysplasia verruciformis (EV) is a genetic disease that leads to an abnormal susceptibility to a large number of β HPV types. Patients with EV present with generalized cutaneous verrucae planae, pityriasis versicolor-like macules, and an early development of squamous cell cancer (SCC) in sun-exposed sites. In the general population, β HPV types are acquired early in life, and they have also been implicated in the initiation, but not maintenance, of SCC in immunocompromised patients.
Keywords
human papillomavirus, HPV, warts, verrucae, condylomata acuminata, anogenital warts, venereal warts, verrucae planae, flat warts, Buschke–Löwenstein tumor, wart treatment, virus-like particles, VLP, HPV vaccine, vulvar intraepithelial lesion (VIN), penile intraepithelial lesion (PIN)
▪ Verrucae vulgares (singular: verruca vulgaris) – common warts ▪ Verrucae palmares et plantares – hand and foot warts ▪ Verrucae planae – flat warts, plane warts ▪ Condylomata acuminata (singular: condyloma acuminatum) – genital warts ▪ Condylomata plana – flat cervical condylomas, plane condylomas ▪ Squamous intraepithelial lesion (SIL) – cervical intraepithelial neoplasia (CIN); vulvar (VIN); vaginal (VaIN); penile (PIN); anal (AIN) ▪ Buschke–Löwenstein tumor – giant condylomata acuminata, condylomata acuminata gigantea ▪ Florid oral papillomatosis – Ackerman tumor ▪ Papillomatosis cutis carcinoides – Gottron tumor ▪ Focal epithelial hyperplasia – Heck disease
- ▪
Human papillomaviruses (HPVs) comprise a large group of more than 150 genotypes that infect the epithelia of skin or mucosa and most commonly cause benign papillomas or warts
- ▪
Epidermodysplasia verruciformis (EV) is an inherited disease characterized by generalized flat warts or pityriasis versicolor-like macules, caused by chronic infection with HPV types in the genus β; patients often develop squamous cell carcinomas (SCCs) in sun-exposed areas
- ▪
EV-associated HPV types are also detected in skin cancers and normal skin. These viruses are activated by UV exposure, immunosuppression, and specific host genetic background (EV)
- ▪
Anogenital HPV is a highly prevalent, sexually transmitted infection (STI) seen predominantly in young adults. Condylomata acuminata or benign anogenital warts are typically caused by HPV-6 or -11, which are considered to be low-risk types
- ▪
Persistent infection with high-risk HPV types, predominantly HPV-16 and -18, is the major cause of cervical and anal cancers and a subset of vaginal, vulvar, penile, oropharyngeal, and rarely digital SCCs
- ▪
Patients with cellular immunodeficiencies are at higher risk for persistent HPV infection and progressive disease
- ▪
No effective specific antiviral treatment for warts exists, and most therapies focus on destruction of visible lesions or induction of a cellular immune response
- ▪
Highly effective prophylactic HPV vaccines based on virus-like particles (VLPs) have been licensed for the prevention of genital warts and/or associated cervical, vulvar, vaginal, and anal cancer
Introduction
Papillomaviruses are a large group of DNA viruses that are widely distributed in animals and humans, most commonly inducing benign papillomas or warts . Recurrent skin and anogenital warts may be disfiguring, impose a considerable psychological burden, and are a frequent cause of medical office visits. A subset of human papillomaviruses (HPVs) designated as high-risk types, most often HPV-16 and -18, is now recognized as the primary etiologic agent for: cervical cancer and its precursor lesions; a subset of the malignancies at other anogenital sites and in the upper aerodigestive tract; and, rarely, squamous cell cancer (SCC) of the digits. In contrast, infection with the common cutaneous HPV types 1, 2, 4, 27, 57, etc. is not thought to have any oncogenic potential . In immunocompromised patients, HPV infections tend to persist and result in an increased risk of developing anogenital neoplasias. The high prevalence of genital HPV infection in sexually active young adults is a major concern, as effective antiviral treatments do not exist. More than two decades ago, prophylactic vaccines based on virus-like particles (VLPs) that prevent transmission in animal models of papillomavirus infection were developed . Human vaccine studies subsequently demonstrated safety and nearly complete (>90%) efficacy in preventing vaccine type-specific genital HPV infection and the development of associated neoplasias .
History
An infectious etiology for human warts had long been suspected , and experimental transmission of common warts with a cell-free extract was demonstrated over 100 years ago . The descriptions of cottontail rabbit papillomavirus (CRPV) in 1933 and of the rare genodermatosis epidermodysplasia verruciformis (EV) demonstrated the oncogenic potential of papillomaviruses. Following the cloning of the first papillomavirus genome in the late 1970s, the plurality of human and animal papillomaviruses became evident . Based on genome sequencing, HPV types can be grouped into phylogenetic trees with a remarkable relationship to biologic behavior, e.g. mucosal and cutaneous types, high-risk and low-risk genital HPV, EV-associated HPV . Today, the genomes of more than 150 HPV types have been fully characterized and additional partial DNA sequences have been obtained, indicating the existence of at least 200 HPV genotypes . In 1976, zur Hausen proposed an association between papillomaviruses and cervical cancer , and subsequent molecular studies led to the recognition of a subset of HPV types in a high proportion of cervical carcinomas. The identification of biologic differences between “high-risk” and “low-risk” mucosal HPV confirmed that infection with oncogenic HPV was the main cause of cervical cancer and its precursor lesions .
Epidemiology
Cutaneous warts are caused by a small group of specific HPV types, with a prevalence of up to 30% in primary schoolchildren and a decline thereafter with increasing age . Patients living in larger households often report an infected cohabitant, supporting the concept of person-to-person transmission. The majority of warts will regress spontaneously within 1–2 years. After clearance occurs, reinfection with the same HPV type appears to be uncommon, suggesting that protective type-specific immunity may develop.
Genital infection with HPV occurs most commonly by intimate contact, while infection of non-genital skin may occur via direct skin-to-skin contact or indirectly through contaminated surfaces and objects (e.g. swimming pool, gymnasium). Basal keratinocytes, which serve as the primary targets for HPV infection, are exposed to the virus through minor abrasions, and infection is promoted by maceration. Autoinoculation of virus from the lesion to adjacent skin is observed frequently, especially for digital or flat warts. Using sensitive polymerase chain reaction (PCR)-based detection, normal-appearing skin surrounding HPV-associated lesions and even skin of healthy volunteers may contain HPV DNA. These observations help to explain the high recurrence rate of warts (e.g. 20–50% for genital condylomata) and the observation that treatment may not prevent further transmission of the virus. Papillomaviruses are resistant to heat and desiccation due to the absence of a viral envelope, and even laser fumes may contain infectious virions .
Genital warts in prepubertal children are uncommon and their diagnosis raises special concerns. Although viral transmission may occur during delivery, from close family contacts, or by autoinoculation from skin warts, the possibility that these lesions may have been caused by sexual abuse should always be carefully considered (see below) .
In the US, HPV infection of the lower anogenital tract is the most common sexually transmitted infection (STI) , with an overall prevalence of ~40% in those ages 19–59 years . However, since introduction of the quadrivalent HPV vaccine, the prevalence of HPV types 6, 11, 16 and 18 in cervicovaginal specimens has decreased in women ages 14–19 years, declining from 11.5% in the prevaccine era (2003–2006) to 5% in the years 2007–2010 . Behavioral risk factors include sexual intercourse at an early age and the number of lifetime sexual partners . Circumcised men are less likely to carry and transmit HPV infection. In men who have sex with men (MSM), anal HPV infection is very prevalent (up to 75%). The public health impact of genital HPV infection is evidenced by the high number of initial visits to physicians’ offices for treatment of genital warts (e.g. 400 000 in 2013 in the US) and the frequency of consequences such as abnormal Pap smears and cervical neoplasia. The annual cost burden of genital HPV infection is estimated to be $6 billion in the US, making it the second most costly STI after HIV infection .
Most genital papillomavirus infections resolve spontaneously, whereupon HPV DNA becomes undetectable by PCR. The median duration of high-risk HPV infections in women is 8 months and persistence is found in 30% after 1 year and in 9% after 2 years . Persistent cervical infection with a high-risk HPV type is required for the development of cervical intraepithelial neoplasia (CIN) and carcinoma. HPV-16 is detected in ~50% of cervical cancers and high-grade CIN. Together, HPV-16, -18, -31, -33, -45, -52 and -58 have been identified in ~90% of cervical cancers .
Worldwide, an estimated 525 000 cases of cervical cancer are diagnosed annually, resulting in about 275 000 deaths . The disease occurs predominantly in low-income countries that lack effective cervical screening programs. In the US, prevention and early detection programs have reduced the incidence rate of cervical cancer by ~75%, to 8 per 100 000 women per year. According to the American Cancer Society, an estimated 12 900 new cases of cervical cancer occurred in 2015, resulting in slightly over 4000 deaths. Although high-risk oncogenic HPV types account for most of the observed risk for developing cervical cancer, smoking, cervical inflammation, parity, and oral contraceptive use have been proposed as cofactors . While there is little evidence for hematogenous spread of HPV, ~60% of patients with anogenital infections develop low-titer, type-specific antibodies during the course of infection . Titers tend to decrease slowly and may persist for years after viral clearance.
Immune suppression in HIV-infected patients and organ transplant recipients results in HPV infections that are more frequent, persistent, and likely to progress to intraepithelial neoplasias, with CD4 + T-cell depletion and HIV viral load being important risk factors. In studies analyzing anal swabs, up to 93% of HIV-seropositive MSM were positive for HPV DNA, compared to 60% of HIV-negative MSM . HPV-associated anal intraepithelial neoplasia (AIN), a likely anal cancer precursor, is found in up to a third of HIV-infected MSM, compared to 20% of HIV-negative MSM. Antiretroviral therapy has shown no or only a modest beneficial effect on the incidence and progression of AIN or CIN. Similarly, organ transplant recipients are at an increased risk for developing warts and genital neoplasias . They also have a higher incidence of premalignant and malignant skin neoplasias, e.g. actinic keratoses, Bowen disease, SCCs, especially in sun-exposed areas.
Cutaneous tumors from organ transplant recipients as well as “healthy skin” and plucked hairs from immunocompetent patients often contain DNA from β HPV types (see below) . Only in patients with EV and immunocompromised hosts do β HPV infections result in highly active viral replication with cytopathic effects and clinical lesions . In skin swab samples, children from 1 month to 4 years of age have a high prevalence (from 50% to 70%) of HPV DNA , indicating that normal-appearing human skin is colonized with a multiplicity of β HPV types very early in life, most likely through close skin-to-skin contact. Beta HPV DNA has also been detected in psoriatic lesions and skin cancers from PUVA-treated patients; however, highly sensitive PCR-based methods were required for detection and viral gene transcription has not been reported.
Recurrent respiratory papillomatosis (RRP) is characterized by exophytic lesions of the airways. It occurs in juvenile- and adult-onset forms and is usually caused by HPV-6 and -11. Despite its low incidence of 0.4–1.2 per 100 000 , RRP is the most common benign tumor of the larynx. It is believed that in childhood-onset RRP, vertical HPV transmission from mother to child occurs by aspiration during delivery through an infected birth canal . Although treatment should be considered in pregnant women with condylomata in order to reduce viral load, there is insufficient evidence to support cesarean section to prevent development of RRP in the offspring. Adult-onset RRP more likely results from genital-to-oral transmission.
Pathogenesis
Papillomavirus Evolution
Historically, papillomaviruses were grouped together with polyomaviruses, but their fundamentally different biology and genetic organization resulted in their reclassification as a separate family in 2000 . The National Center for Biotechnology Information (NCBI) nucleotide sequence database ( www.ncbi.nlm.nih.gov ) currently lists more than 150 different HPV types as well as numerous incompletely characterized putative new HPV types. By definition, a novel type has more than 10% dissimilarity from any previously characterized type in the DNA sequences of three HPV genes (E6, E7, and L1). A dissimilarity of 2–10% is defined as a subtype, and <2% as a variant .
By comparing DNA sequences in conserved portions of the papillomavirus genomes across different types, a phylogenetic tree establishing the degree of relatedness among these types has been generated . Two genera, the α- and β-papillomaviruses, encompass almost all known HPV types, with the genus α containing mucosal and cutaneous HPV types that are pathogenic in normal hosts and the genus β containing the types associated with EV; γ, µ, and ν genera contain additional cutaneous types. Within each genus, HPV types are grouped according to sequence homology into species, which often share similar biologic and pathologic properties (e.g. HPV-16 and -31). Present models postulate that HPVs co-evolved with our human ancestors and novel HPV types or variants were generated through either genetic drift or natural selection via adoption of specialized ecologic niches. Genetic recombination between HPV types does not occur.
Virology
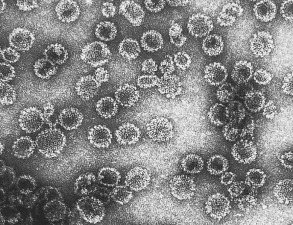
Papillomaviruses are non-enveloped, double-stranded DNA viruses approximately 55–60 nm in diameter . The spherical capsid is composed of two virally encoded proteins: the major structural protein L1 and the minor structural protein L2. During virion assembly within cells, L1 proteins form pentamers called capsomeres, and 72 capsomeres multimerize to form the viral capsid with an icosahedral symmetry arranged on a T = 7 surface lattice. The capsid surrounds the viral DNA, thereby protecting it from degradation, and it also enables the virus to bind efficiently to target cells. Packaged viral DNA is associated with L2 and cellular histones, forming a mini chromosome. In contrast, the early (E) viral proteins are not incorporated into infectious virions.
When expressed in cell culture, the L1 major capsid proteins of papillomaviruses self-assemble into VLPs that are morphologically similar to native virions ( Fig. 79.1 ) . Papillomavirus VLPs display type-specific and neutralization epitopes that are used to detect serum antibodies and as the basis for prophylactic vaccines (see below) .
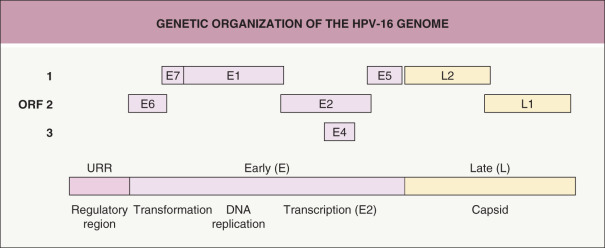
The HPV genome, ~8 kilobases (kb) in length, is composed of three domains: the upstream regulatory region (URR), the early region, and the late region ( Fig. 79.2 ). The URR, ~1 kb in length, lacks open reading frames (ORFs) and contains the origin of replication and many of the control elements for transcription and replication. The early region, ~4 kb in length, contains the ORFs for genes expressed early in the papillomavirus life cycle. The late region, ~3 kb in length, encodes the capsid proteins (see above) .
Papillomavirus Life Cycle
The papillomaviruses are highly species-specific, and productive infection has never been observed outside natural host tissue. The papillomavirus life cycle is completed only in fully differentiated squamous epithelia. This has impeded its study in monolayer tissue culture cells, where late gene expression and virion production do not occur. Only a small number of HPV types have been successfully propagated in mouse xenograft or raft culture systems, and limited amounts of infectious virions have been generated .
Productive infection and induction of hyperproliferation are initiated when the virus enters proliferating basal epithelial cells ( Fig. 79.3 ). This layer of cells is not normally accessible to the virus because of the mechanical barrier provided by the overlying differentiated cell layers. Therefore, it is likely that infection requires an abrasion or other trauma to the epithelium to expose the basal cells to the virus. Our knowledge about early steps of infection such as viral entry and uncoating is limited. The receptor(s) that mediate virus binding to epithelial cells have not been definitively identified, but binding appears to depend on the L1 major capsid protein, and cell surface and/or basement membrane heparan sulfate is necessary for efficient infection in vitro . The amino terminus of the L2 minor capsid protein is then cleaved by furin (a proprotein convertase), which exposes both a cross-neutralization epitope on L2 and a binding site (thought to be on L1) that facilitates viral entry via a putative secondary receptor on keratinocytes . Upon infection, the covalently closed circular DNA genome becomes established as a low-copy autonomous replicon in the nuclei of basal cells, thereby creating a long-term reservoir of viral DNA.
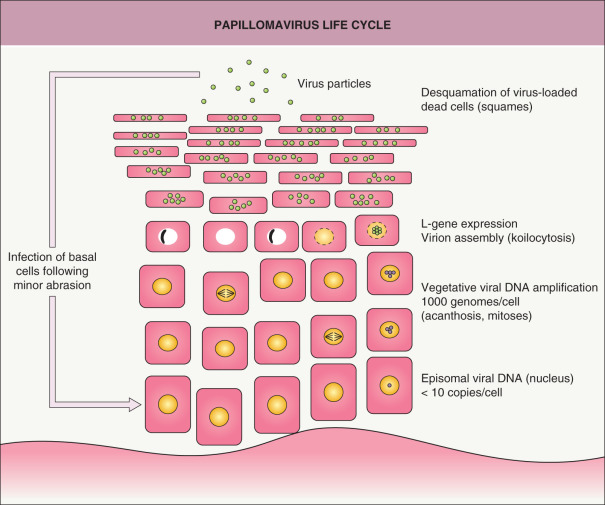
The program of viral gene expression is intimately linked to the differentiation state of the infected cells. The E genes are transcribed at low levels in the basal cell layer and spinous layer. The first genes to be expressed following infection are the E1 and E2 genes, which are responsible for controlling the transcription of other viral genes and replication of the viral genome. HPVs do not encode the enzymes required for transcription or replication of viral DNA. Consequently, HPV is entirely dependent on the co-opting of cellular machinery for these functions. One major effect of HPV proteins E5, E6 and E7 is that the epidermal cell cycle, which is normally blocked for cells that are suprabasal, continues so that HPV genome copy number can be amplified to high levels during vegetative viral replication for assembly into virions (see Fig. 79.3 ). The E6 and E7 proteins of the high-risk mucosal HPV types act as viral oncoproteins, but no such functions are associated with the corresponding proteins of the low-risk mucosal and EV types . There is limited understanding of possible differences between oncogenic (e.g. HPV-5, -8) and the non-oncogenic β HPV types.
In combination with other cellular proteins, E6 from high-risk mucosal HPV causes the ubiquitin-mediated degradation of the cellular protein p53. Elevated levels of p53 arrest cells in the G1 phase of the cell cycle or induce apoptotic cell death, so that E6-facilitated destruction of p53 removes a brake on suprabasal cell cycling. One of the principal effects of high-risk mucosal HPV E7 is to bind to the underphosphorylated form of RB (the retinoblastoma tumor suppressor protein). A physiologic role of underphosphorylated RB is to bind to and inhibit the function of the E2F transcription factor (see Ch. 107 ). When HPV E7 binds RB, E2F is liberated from this inhibition and is thereby able to induce the expression of genes required for DNA replication. Both E6 and E7 are multifunctional proteins, and while their effects on p53 and RB are critical ones, they do have additional targets important to the oncogenic potential of the virus. These include activation of telomerase by E6, binding of histone deacetylases by E7, and synergistic effects of both proteins that lead to centrosome abnormalities and chromosomal instability .
The genes for the virion structural proteins, i.e. the late (or L) genes, are expressed in the terminally differentiated superficial layers of the epithelium. The more superficial epithelial layers also have higher levels of expression of E1 and E2 and amplification of viral DNA. The amplified genomes are encapsulated by the L1 and L2 capsid proteins to generate infectious virions, and virus particles are observed in the granular layer of the epithelium and above. Virus assembly is not believed to lyse the cells; instead, the virus is shed with the cornified layer as cells slough from the epithelial surface (see Fig. 79.3 ). E4 proteins are hypothesized to disrupt the intracellular filamentous network of keratinocytes, which may facilitate virus release from the corneocytes .
Host Immune Response
Persistent papillomavirus infections are common, indicating that HPVs have evolved mechanisms to evade immune surveillance. There is no viremic phase during the life cycle, so a systemic immune response is avoided. In addition, low levels of viral proteins are expressed in the basal and spinous cell layers of the epidermis, where they would be most likely to be recognized by Langerhans cells and infiltrating lymphocytes. Only in the more immunologically privileged terminally differentiated layers is there extensive production of virion proteins, with shedding of infectious virus exclusively from the external epithelial surface . Despite the relative success of papillomaviruses in evading immune responses, up to two-thirds of cutaneous warts spontaneously regress within 2 years, and lesions of multifocal infections often regress concomitantly. There is also an increased prevalence of warts in patients with suppressed cell-mediated immunity. In contrast, patients with underlying defects in humoral immunity do not seem to be predisposed to HPV infections, suggesting a primary role of cell-mediated responses.
Although low amounts of virion capsid proteins are produced in the lower epidermal cell layers, serum antibodies specific to conformational virion epitopes of high-risk HPV-16 can be detected in more than half of women with low-grade or asymptomatic genital HPV-16 infections . It is unknown if these antibodies play a role in limiting reinfection with the same type from autoinoculation or from an infected sexual partner.
Latent HPV infections are defined as persistence of HPV DNA in clinically and cytologically normal epithelium. However, it is difficult to distinguish true viral latency from persistent infection with a low level of DNA replication. The mechanisms by which HPV enters into and is activated out of latency are unknown .
Oncogenic Potential of HPV
Different HPV types have markedly different oncogenic potentials. The vast majority of cervical cancers, for example, contain the sequences of one of five HPV types: 16, 18, 31, 33 or 45 . In contrast, infection with HPV-6 or -11 occurs commonly in benign or low-grade intraepithelial lesions but is very rarely associated with the development of anogenital malignancies. HPV-5 and -8 DNA sequences are frequently detected in SCCs associated with EV.
Progression of HPV-associated cervical lesions to invasive cancer typically requires several decades. Both persistent HPV infection and the accumulation of additional genetic mutations are required in the current model of multistage carcinogenesis . Integration of the viral genome randomly into the host DNA is a common event, invariably leading to loss of E1 and E2 expression. In HPV-16 and -18, E2 functions to repress the transcription of E6 and E7 genes. Therefore, integration is thought to increase expression of E6 and E7, which are selectively retained and expressed at elevated levels in carcinomas, suggesting a critical role for oncogenic progression (see above) .
Differences in the activities of E6 and E7 genes between oncogenic and non-oncogenic HPV types principally account for the different risk of progression. E6 and E7 proteins derived from high-risk HPV types (e.g. 16 and 18) degrade p53 and interact with RB (respectively) with greater efficiency than do E6 and E7 proteins from low-risk types (e.g. 6 and 11) . Degradation of p53 by E6, which eliminates from infected cells the principal “guardian of the genome” (the protein responsible for cell cycle arrest in the presence of damage to cellular DNA), can lead to the accumulation of genetic mutations in infected cells. Thus, carcinogenic progression appears to be promoted or facilitated by increased E6 and E7 expression as well as by activity of external carcinogens, resulting in genomic instability .
Clinical Features
Cutaneous and mucosal HPV types form two distinct groups that infect either skin or mucosa. However, viral tropism is not absolute, as DNA of skin types may be found in genital or oral warts, and in rare cases, mucosal types may be associated with skin lesions. Clinical manifestations depend on the HPV type involved, the anatomic location, and the immune status of the host ( Table 79.1 ).
| CLINICAL MANIFESTATIONS AND ASSOCIATED HUMAN PAPILLOMAVIRUS (HPV) TYPES | ||
|---|---|---|
| Frequently detected | Less frequently detected | |
| Skin lesions | ||
| 1, 2, 27, 57 | 4, 29, 41, 60, 63, 65 |
| 3, 10 | 28, 29 |
| 7 | 1, 2, 3, 4, 10, 28 |
| 16 | 26, 31, 33, 34, 35, 51, 52, 56, 73 |
| 3, 5 , 8 | 9 , 12 , 14 , 15 , 17 , 19–25 , 36–38 , 47 , 49 , 50, etc. |
| 5 , 8 | 14 , 17 , 20 , 47 |
| Mucosal lesions | ||
| 6, 11 | 40, 42–44, 54, 61, 70, 72, 81 |
| 16 | 18, 26 * , 31, 33, 35, 39, 45, 51, 52, 53 * , 56, 58, 59, 62, 66 * , 68, 73, 82 |
| 6, 11 | |
| 6, 11 | |
| 13, 32 | |
Cutaneous Infections
Cutaneous HPV types comprise a small group of viruses that infect the skin and induce common warts (Latin verrucae vulgares ; singular verruca vulgaris ), palmar and plantar warts ( verrucae palmares et plantares ), mosaic warts, flat warts ( verrucae planae ), and butcher’s warts (see Table 79.1 ). In general, classification of warts is based on morphology, histology, and anatomic location.
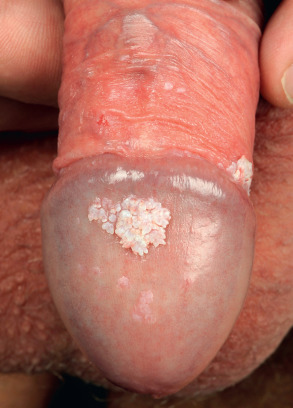
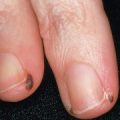
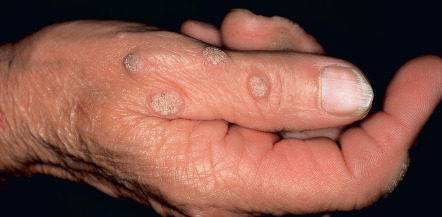
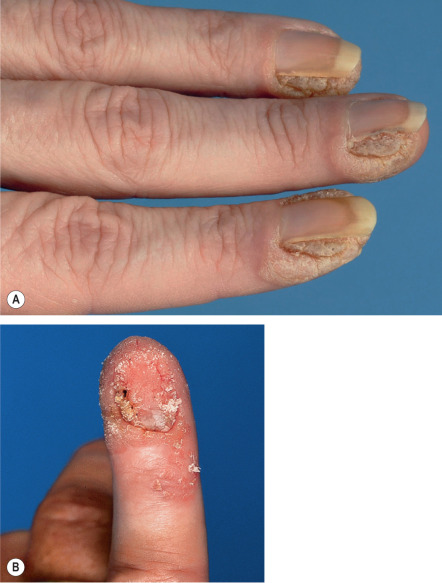
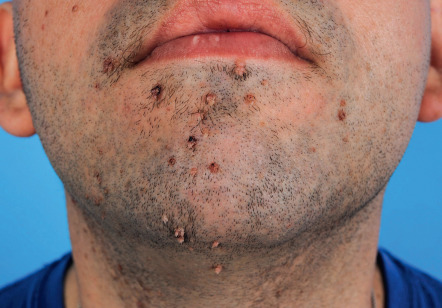
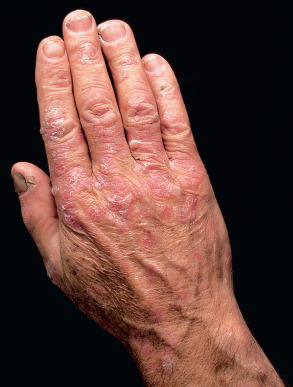
Common warts are hyperkeratotic, exophytic, dome-shaped papules or plaques that are typically associated with HPV-1, -2, -4, -27, or -57. These warts are most frequently located on the fingers and dorsal surfaces of the hands ( Figs 79.4 & 79.5 ) or in other sites prone to trauma such as the knees or elbows, but they may occur anywhere on the skin surface. Involvement of the proximal nail fold and/or ablative therapy at that location may destroy the matrix, resulting in onychodystrophy (see Fig. 79.5 ). Characteristic features of common warts are punctate black dots representing hemorrhage into the stratum corneum ( Fig. 79.6 ). Autoinoculation by scratching may cause a linear arrangement of warts. Slender, exophytic filiform warts can also develop, especially in periorificial locations on the face ( Fig. 79.7 ).
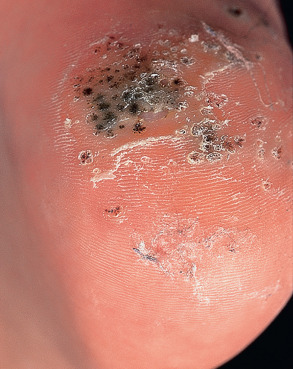
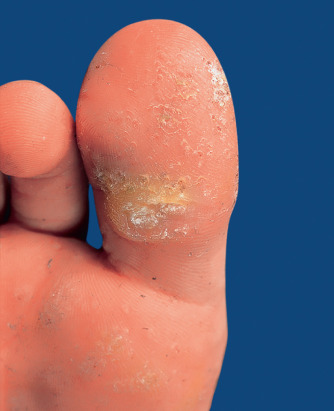
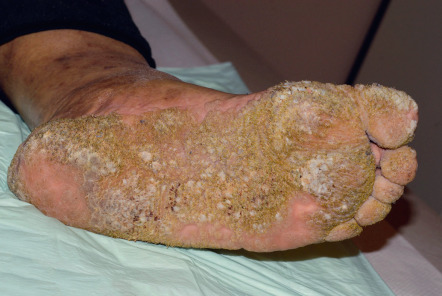
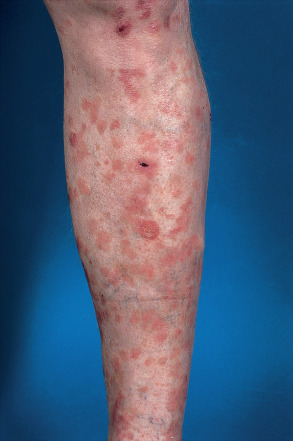
Palmar and plantar warts appear as thick, endophytic papules on the palms, soles, and lateral aspects of the hands and feet, with gently sloping sides and a central depression resembling an anthill (hence the term myrmecia , meaning anthill). On the soles, these are often painful from pressure when walking, due to their deep inward growth ( Fig. 79.8 ). Plantar warts that coalesce into large plaques are referred to as mosaic warts (see Fig. 79.8 ). Extensive chronic verrucosis that is notoriously resistant to therapy has been observed in immunocompromised individuals and, at times, in patients with no apparent immune dysfunction ( Fig. 79.9 ). Inclusion warts of the sole are plantar cysts from which HPV types 4, 60, 63, and 65 have been isolated . In studies from Europe, HPV-1 and the closely related types HPV-2, -27, and -57 caused the majority of palmoplantar warts, and they occurred most commonly in patients 6–10 years of age . Although a higher incidence of HPV-2-related warts was reported in atopic children, other investigators have failed to find such an association.
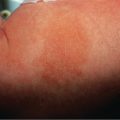
Flat warts are skin-colored or pinkish to brown, relatively smooth-surfaced, slightly elevated, flat-topped papules that are most commonly located on the dorsal hands, arms or face, often in a linear array ( Fig. 79.10 ). They are usually caused by HPV-3 or -10, and less often by HPV-28 and -29.
Butcher’s warts , which earn their name from their occurrence in meat- (or fish-) processing professionals, appear as extensive verrucous papules or cauliflower-like lesions on the dorsal, palmar, or periungual aspects of the hands and fingers. These warts are associated with HPV-7 not animal papillomaviruses.
Epidermodysplasia verruciformis (EV), first described in 1922 by Lewandowski and Lutz, is a rare genetic disease. It is characterized by a particular susceptibility to cutaneous infections with HPV types in the genus β (see above), which do not produce clinical lesions in immunocompetent individuals . The disease usually manifests in childhood with highly polymorphic, widespread lesions. About two-dozen specific HPV types have been described in EV patients, a subset of which (mainly types 5 and 8) is also detected in EV-associated skin cancers. EV typically has an autosomal recessive pattern of inheritance and is caused by truncating mutations in two genes, TMC6 (EVER1) and TMC8 (EVER2) , that encode transmembrane proteins localized to the endoplasmic reticulum . These proteins form a complex that interacts with the zinc transporter 1 (ZnT1) and affects intracellular zinc distribution, down-regulating zinc-associated transcription factors . Inhibition of ZnT1-TMC6/8 function by the HPV E5 protein is thought to facilitate viral infection; β HPV types lack E5, but they become pathogenic in EV patients with TMC6/8 defects .
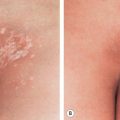
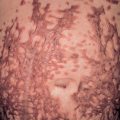
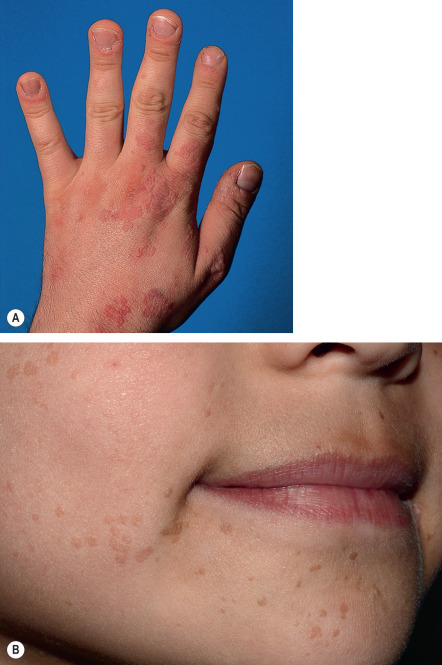
Patients with EV present with widespread, discrete or confluent papules that resemble flat warts ( Fig. 79.11 ); scaly, pinkish or hypopigmented guttate macules and thin plaques that resemble pityriasis versicolor represent another characteristic finding ( Fig. 79.12 ). In an individual patient with EV, warts that appear clinically distinct often contain different HPV types, and each skin lesion usually contains several HPV types. Actinic keratoses usually arise after the age of 30 years and slowly transform into invasive SCCs in approximately half of EV patients. These tumors have low metastatic potential and develop primarily in sun-exposed areas such as the forehead, ears and hands, implicating UV irradiation as an important co-carcinogen. Patients with EV should be educated about the importance of minimizing sun exposure, and family members should be screened for the presence of similar lesions.
It may be difficult to differentiate flat warts from EV-associated lesions based on histology alone (see below), but the presence of lesional β HPV DNA (e.g. HPV-5, -8, etc.) can establish the diagnosis in an otherwise healthy individual. Beta HPV types can also induce a sporadic EV-like eruption in immunosuppressed patients, including transplant recipients and those with AIDS or a lymphoma. In addition, β HPV sequences have been isolated from the normal skin of healthy individuals by highly sensitive DNA detection methods, indicating that the general population acts as the reservoir for these HPV types. There is increasing epidemiologic and biologic evidence that β HPV types may play an adjunct role in the initiation, but not maintenance, of SCCs in immunosuppressed patients .
WHIM syndrome is a rare, autosomal dominant primary immunodeficiency disorder characterized by HPV-induced w arts (cutaneous and genital), h ypogammaglobulinemia, recurrent bacterial i nfections, and neutropenia due to m yelokathexis (retention of mature neutrophils in the bone marrow) (see Ch. 60 ). It is caused by mutations in the gene encoding the chemokine CXC receptor 4 (CXCR4), which result in impaired leukocyte trafficking and homing .
WILD syndrome is a more recently characterized disorder that features w arts, cellular i mmunodeficiency, primary l ymphedema and multifocal anogenital d ysplasia. The patient reported had extensive warts, including condylomata acuminata and lesions that clinically (but not histologically) resembled those in EV. A variety of α HPV types were detected in the cutaneous and genital lesions .
Several other primary immunodeficiencies are associated with severe HPV infections, including DOCK8 deficiency, GATA2 deficiency (MonoMAC), and idiopathic CD4 + lymphopenia .
Mucosal Infections
More than 40 HPV types preferentially infect the mucosa of the anogenital and upper aerodigestive tracts. Subclinical infections are much more common than visible warts. Application of 5% acetic acid (aceto-whitening) may aid in the identification of subclinical lesions as white areas.
Condylomata acuminata (singular: condyloma acuminatum ), or anogenital warts, are found on the external genitalia and the perineum, perianally, or in adjacent areas such as the inguinal fold and mons pubis. Lesions may extend into the vagina, urethra, or anal canal (but rarely beyond the dentate line). Condylomata are typically discrete, sessile, smooth-surfaced exophytic papillomas or acuminate warts and may be skin-colored, brown or whitish (especially when macerated in moist areas) ( Fig. 79.13 ). They lack the thick cuirass of horny scale present on cutaneous warts and frequently measure one to several millimeters in diameter. Condylomata may also present as pedunculated or broad-based papillomas up to several centimeters in diameter or as large confluent plaques ( Fig. 79.14 ). Condylomata plana , or flat cervical warts, may be difficult to recognize without aceto-whitening and the use of a (magnifying) colposcope. High-grade intraepithelial neoplasias are usually caused by high-risk types, mainly HPV-16, -18 and -31, whereas low-grade lesions may contain low-risk as well as high-risk HPV types.