Chapter 3 THE NICHE THEORY FOR FAT GRAFT SURVIVAL
The biologic mechanisms responsible for the survival of grafted fat are still not completely understood, despite decades of study. On the basis of the most recent studies performed on a large number of human specimens of adipose tissue, it is possible to provide additional information about the processes involved in the survival of grafted fat and volume maintenance after fat grafting. 1 The unresolved problem is still maintaining the volume of grafted adipose tissue, in particular after breast reconstruction and for facial defect correction, where the proportions of volumes are most critical.
The discovery of cell populations with stem properties within adipose tissue have prompted reconsideration of the potential clinical applications of fat. 2 In the field of plastic and reconstructive surgery, the traditional concept of fat grafting is evolving toward the new concept of liporegeneration. 3 Adipose tissue should be considered not as a filler, but as the source of regenerative units. Until the discovery of the stem properties of adipose-derived stromal cells (ADSCs), the favorable outcome of fat transplants was simply attributed to the capacity of fat cells to survive and preserve their morphologic features after heterotopic transplantation. However, the yield of surviving cells proved controversial, leading to debates about the importance of endogenous and exogenous adipocytes for graft outcome. 4 The viability of transplanted fat cells has also been doubted, giving rise to the question of whether the success of a graft is ultimately the result of surviving grafted adipocytes (the cell survival theory) or the recruitment of host adipocytes (the host replacement theory). 5
Today this debate has evolved into an inquiry about the role of ADSCs, their survival, and the extent and modality of their involvement in regenerative processes. To answer these questions, we must also consider the complex relationships that ADSCs establish with all the external factors, physiologic (before transplant) or ectopic (after transplant), that contribute to determining their identity and fate—that is, the niche.
In this chapter we will discuss the concept explaining fat graft survival on the basis of niche theory. We will provide a general definition of niche and explain how niche theory can support crucial advances in stem cell biology and help to optimize the new, extraordinary therapeutic potential of grafted fat.
The Stem Cell Niche of Fat
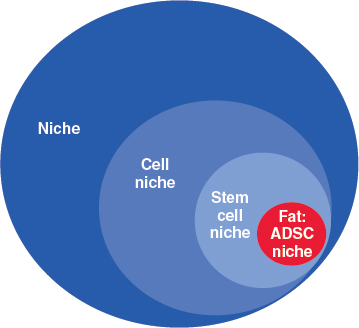
Through a comprehensive discussion of the definition of niche and stem cell niche, we will introduce the concept of fat as the niche of adipose-derived stromal cells.
THE NICHE
The Word Web Online Dictionary provides the following four definitions for niche: (1) a position particularly well suited to the person who occupies it; (2) a small concavity; (3) an enclosure that is set back or indented; and (4) (ecology) the status of an organism within its environment and community (affecting its survival as a species). A fifth meaning can be derived from economics, where the term “niche market” (also known as “target market”) refers to a specific targetable portion (subset) of the market.
In a physical space, a niche is a recess, a restricted portion of that space, that is usually associated with an idea of suitability and that allows whoever or whatever occupies it to satisfy his needs and express properties inherent in his identity. Within complex systems such as an ecosystem or a market, a niche includes the range of elements and interactions that allow the long-term survival of a given species or business, respectively.
The Cell Niche
The physiology of any cell is strictly regulated by the interactions that it establishes with its microenvironment (niche). These interactions involve physical communications with neighboring cells or the surrounding extracellular matrix and the binding of circulating or paracrine factors released by distant or local cells, respectively. In other words, the niche concept, which originally referred to interactions between species and their environment in the context of the whole ecosystem, can be extended to describe the complex relationships inside a single organism that allow any cell to survive, grow, differentiate, and reproduce.
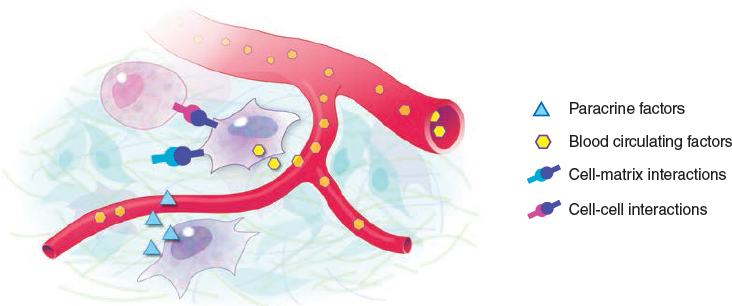
The cell niche concept applies to interactions with neighboring cells or the surrounding extracellular matrix, but also to long-distance interactions through the blood circulation, lymphatic system, and nerve pathways. Thus the phenotypic identity of any single cell and its reaction to a given stimulus are not determined only by the genetic or epigenetic equipment of the cell, but are also strongly dependent on the specific niche context in which the cell resides. A change in niche parameters, or transplantation of cells into niches other than their original ones, can drastically affect cell physiology and alter its properties.
The cell niche theory can obviously be applied to any kind of cell, but the impact of this concept is particularly significant in the context of stem cell biology. The characteristic that primarily distinguishes stem cells from differentiated cells is their plasticity, or capacity to change their identity in relation to their external environment, or niche. Changes in the niche of a stem cell might plausibly have a profound impact on the plasticity of the stem cell, resulting in its arrest, through cell commitment; its activation toward a regenerative or reparative function; or its aberrant evolution toward neoplastic behavior. Considering the same concept in terms of clinical applications, the outcome of therapeutically aimed cell grafting into an exogenous environment could be understood, predicted, and controlled only on the basis of knowledge, or prediction, of the local and distant factors that govern the physiology of grafted cells and their responsiveness to incoming stimuli.
The cell niche theory is only in its infancy, and our preliminary considerations are obviously simplistic. In this chapter we will discuss recent insights into different examples of stem cell niches, with the ambitious aim of defining common biologic paradigms that could help us to better understand and predict the outcome of therapeutic grafting of the cells we are interested in—ADSCs—along with their niche—fat.
Stem Cell Niches
The concept of a niche as a specialized microenvironment housing stem cells was first proposed by Schofield 6 almost 40 years ago with reference to mammalian hematology. Although the field of stem cell biology is still young and very confusing, several aspects have been clarified for certain stem populations, providing reference concepts with the potential to define a common paradigm of the stem cell niche.
The first well-known stem cell niche is that of hematopoietic stem cells (HSCs). HSCs are mainly found in bone marrow and, to a lesser extent, in extramedullary tissues, such as the spleen and liver. In bone marrow, HSCs predominantly line the endosteum. Both osteoblasts and osteoclasts have been shown to be involved in the determination of HSC fate. Endosteal osteoblasts are thought to provide a variety of factors that regulate HSC proliferation and contribute to the homeostatic maintenance of the HSC reservoir, 7 such as osteopontin, KIT ligand, and Notch ligand (Jagged-1). It is possible to observe how different environmental factors can influence the signaling of the HSC niche.
In bone marrow, as well as in the spleen and liver, HSCs can also be found adjacent to sinusoids, which points to the perivascular area of sinusoids as another HSC niche. The connection of HSCs with sinusoids could ensure homeostatic blood cell production and prompt responses to hematologic stresses. It has been suggested that endosteal niches maintain HSC quiescence over the long term, whereas perivascular niches maintain HSCs over shorter periods, supporting fate determination and circulation.
Neural stem cells (NSCs) are cells with the molecular and morphologic features of astrocytes that reside in the subventricular zone (SVZ) of the lateral ventricle wall and the subgranular zone (SGZ) of the dentate gyrus of the hippocampus. In addition to the NSCs (type B cells), the neural niche contains rapidly dividing (transit amplifying) cell populations, called type C cells in the SVZ and type D cells in the SGZ, lineage-committed (postmitotic) migratory neuroblasts (type A cells), glial cells, blood vessels, and vessel-associated cytotypes (pericytes, fibroblasts, smooth muscle cells, and macrophages). A large number of molecules are known to modulate the balance between NSC homeostasis and cell fate determination, such as growth factors (noggin, bone morphogenetic proteins [BMP], epidermal growth factor, fibroblast growth factor, vascular endothelial growth factor, pigment-epithelium–derived factor, platelet-derived growth factor), and neurotransmitters (catecholamine, serotonin, gamma-aminobutyric acid [GABA], glutamate, and nitric oxide). 8
Recently an important role in NSC niche homeostasis has been recognized for a highly organized extravascular basal lamina, which is regularly arranged along the subependymal layer and consists of extensively branched stems terminating in bulbs immediately beneath the ependyma. All the cytotypes composing the NSC niche are in direct contact with one or more branches of this basal lamina, suggesting direct or indirect involvement of cell-matrix interactions in NSC fate regulation. 9
The precursors of enterocytes and goblet cells are located in apposite niches within the intestinal crypts. The niches consist of intestinal stem cells (ISCs), transit-amplifying (TA) cells, and pericryptal fibroblasts, so they are made up of an endodermal derived epithelium surrounded by cells of mesodermal origin. Cell signaling between these two tissue layers plays a critical role in coordinating patterning and organogenesis of the gut and its derivatives. The table below summarizes for this stem cell niche the signaling occurring between stem cells and the environment.
Other organ- or tissue-specific stem cell niches (hair follicle, rodent incisor, dental pulp, kidney, liver, and lung) have been described in recent years. Although the picture is still somewhat confused and several aspects are controversial, some substantial analogies can be found between all the various stem cell niches, despite evident differences in cytoarchitecture and histologic features. In particular, the following elements seem to recur as universal regulators of stem cell homeostasis and fate determination: (1) canonical Wnt pathway, (2) cell-matrix interaction, (3) neurotransmitters, (4) interaction with blood vessels, and (5) interaction with cells of mesodermal origin.
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


