Type VII collagen is a major component of the anchoring fibrils of the dermal-epidermal adhesion on the dermal side at the lamina densa/papillary dermis interface. Dystrophic epidermolysis bullosa (DEB) emerged as a candidate for type VII collagen mutations becausing anchoring fibrils were shown to be morphologically altered, reduced in number, or completely absent in patients with different forms of DEB. Circulating autoantibodies recognize type VII collagen epitopes in epidermolysis bullosa acquisita. The suggestion that type VII collagen is required for human epidermal tumorigenesis relates to the increasing numbers of life-threatening complications associated with developing squamous cell carcinomas because of the extended life span of affected individuals with recessive DEB.
The collagen family of proteins
Collagens, the major extracellular matrix components in most vertebrate tissues, comprise a superfamily of proteins. A total of 29 genetically distinct collagens have been described so far in vertebrate tissues and designated by Roman numerals I to XXIX in order of their discovery. The collagen molecules consist of 3 subunit polypeptides, so-called α-chains, and whereas some collagens are homotrimers, others can be heterotrimers containing 2, or even 3, genetically distinct subunit polypeptides. Consequently, there are well over 40 genes in vertebrate tissues that encode the subunits polypeptides of different, genetically distinct collagen molecules.
A characteristic structural feature of all collagens is the presence of a protein domain in triple-helical conformation that provides stability to these molecules to serve as structural building blocks providing integrity to connective tissues. The triple-helical conformation resists nonspecific proteolysis, such as digestion with pepsin. The folding of the individual α-chains into the triple-helical conformation is predicated on the characteristic primary sequence, consisting of repeating Gly-X-Y triplet sequences. In some collagens, such as in type I collagen, the most abundant collagen in the skin and bones, the central collagenous domain of individual α-chains, contains an uninterrupted Gly-X-Y repeat segment spanning approximately 1000 amino acids. In some collagens, such as in type IV (the basement membrane collagen) and type VII (the anchoring fibril collagen), the Gly-X-Y repeat sequence contains imperfections that interrupt the triple-helical conformation. These interruptions then provide flexibility to the rodlike collagen molecules and also provide sites susceptible to nonspecific proteolytic cleavage of the primary sequence.
On the basis of their fiber architecture in tissues, the genetically distinct collagens have been divided into different subgroups. Collagens types I, II, III, V, and X align into large extracellular fibrils and are designated as fibril-forming collagens. Type IV collagen is arranged in an interlacing network within the basement membranes, whereas type VI collagen forms distinct microfibrils and type VII forms anchoring fibrils. Fibril-associated collagens with interrupted triple helices (FACIT) collagens include types IX, XII, XIV, XIX, XX, and XXI. Several of the latter types of collagens associate with larger collagen fibers and serve as molecular bridges, stabilizing the organization of the extracellular matrix.
The major collagens in human skin are types I and III, which account for approximately 80% and 10% of the total bulk of collagen, respectively ( Table 1 ). These 2 collagens associate to form broad extracellular fibers characteristic of human dermis. Type V collagen is present in most connective tissues, including the dermis where it represents less than 5% of the total collagen. In the dermis, type V collagen is located on the surface of the large collagen fibers formed by type I and III collagens, and type V collagen regulates the lateral growth of these fibers. Another major collagen in the skin is type IV collagen, present within the dermal-epidermal junction and in the vascular basement membranes.
| Collagen Type | Chain Composition | Supramolecular Assembly | Tissue Distribution b |
|---|---|---|---|
| I | [α1(I)] 2 α2(I) | Fibrillar | Dermis, bone, tendons |
| III | [α1(III)] 3 | Fibrillar | Fetal dermis, blood vessels, GI tract |
| IV | [α1(IV)] 2 α2(IV) a | Basement membrane | Ubiquitous |
| V | [α1(V)] 2 α2(V) a | Fibrillar | Ubiquitous |
| VI | α1(VI)α2(VI)α3(VI) a | Microfibrils | Ubiquitous |
| VII | [α1(VII)] 3 | Anchoring fibrils | Anchoring fibrils (see table 2 ) |
| VIII | [α1(VIII)] 3 | Network forming | Endothelia |
| XIII | [α1(XIII)] 3 | Transmembrane | Ubiquitous, including epidermis |
| XIV | [α1(XIV)] 3 | FACIT | Skin, cornea |
| XV | [α1(XV)] 3 | Basement membrane | Ubiquitous |
| XVII | [α1(XVII)] 3 | Transmembrane | Hemidesmosomes in skin, cornea, mucous membrane |
| XXIX | Unknown | Unknown | Epidermis |
a Additional α-chains have been identified.
b Distribution in the skin and other major tissues is indicated; lesser amounts may be present in other tissues.
In addition to these major collagens, human skin contains several minor collagens that demonstrate spatially restricted location, yet they play a critical role in providing integral stability to the skin (see Table 1 ). One of them is type VII collagen, the major, if not exclusive, component of anchoring fibrils. Another one is type XVII collagen, a transmembrane collagen in type II topography. Type XVII collagen resides in hemidesmosomes complexed with α6β4 integrin, plectin, and laminin-332 (laminin-5). Finally, type XXIX collagen has been recently reported to be a putative epidermal collagen with the highest level of expression in suprabasal layers. This collagen has been suggested to play a role in atopic dermatitis but its characterization is currently incomplete.
The biology of type VII collagen
Type VII collagen was initially described as an extended, unusually long molecule, hence the original designation as long-chain (LC) collagen. Rotary shadowing electron microscopy of type VII collagen molecules synthesized and secreted by human keratinocytes in culture revealed a long, 424-nm, triple-helical domain and flanking noncollagenous sequences ( Fig. 1 A). The amino-terminal domain was particularly noticeable, with individual α-chains contributing an extended arm. Simultaneously, visualization of type VII collagen isolated by limited pepsin proteolysis of amniotic membranes revealed a 780-nm dimer of 2 identical molecules in antiparallel orientation, with a 60-nm overlap stabilized by disulfide bonds ( Fig. 1 B, C). Further proteolytic digestion with pepsin revealed that type VII collagen molecules consist of a central collagenous, triple-helical segment flanked by the noncollagenous NC-1 and NC-2 domains. Subsequent cloning of the human type VII collagen gene and the corresponding complementary DNA (cDNA) indicated that the initially synthesized type VII collagen subunit polypeptide, the pro-α1(VII) chain is a complex modular protein consisting of a central, 1530-amino acid triple-helical domain ( Fig. 2 A). However, unlike interstitial collagens, the repeating Gly-X-Y sequence is interrupted by 19 imperfections because of insertions or deletions of amino acids in the Gly-X-Y repeat sequence. In the middle of the triple-helical domain, there is a 39-amino acid noncollagenous “hinge” region that is susceptible to proteolytic digestion with pepsin. The amino-terminal NC-1 domain of type VII collagen (NC-1[VII]), approximately 145 kDa in size, consists of submodules with homology to known adhesive proteins, including segments with homology to cartilage matrix protein, 9 consecutive fibronectin type III-like (FN-III) domains, a segment with homology to the A domain of von Willebrand factor, and a short cysteine and proline-rich region. The carboxy-terminal noncollagenous domain, NC-2, is fairly small, approximately 30 kDa, and it contains a segment with homology to the Kunitz protease inhibitor molecule (see Fig. 2 A).
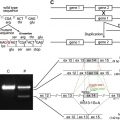
Cloning of the human type VII collagen gene, COL7A1 , revealed a complex structure consisting of a total of 118 separate exons ( Fig. 2 B). However, the gene is fairly compact, and most of the intervening sequences (introns) are fairly small; consequently, the size of the human COL7A1 gene is only approximately 32 kb, encoding a messenger RNA (mRNA) of approximately 8.9 kb. COL7A1 has been mapped to the short arm of human chromosome 3, region 3p21.1. At the time of the report of its structural organization, the type VII collagen gene was noted to be composed of more exons than any previously characterized gene. Most of the COL7A1 introns are small, including a 71-nucleotide intron that was the smallest intron yet reported in a collagen gene. The human type VII collagen gene structure and the encoded primary sequence of the protein are well conserved, and for example, the mouse gene shows 84.7% homology at the nucleotide and 90.4% identity at the protein level, attesting to the importance of type VII collagen as a structural protein.
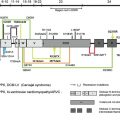
Type VII collagen gene expression displays a restricted, tissue-specific pattern. Specifically, type VII collagen has been localized by immunomapping to a select number of epithelia, including human skin, and the presence of type VII collagen correlated with the presence of ultrastructurally detected anchoring fibrils ( Table 2 ). The expression of the type VII collagen gene can be modulated by several cytokines, and in particular, transforming growth factor-β is a powerful upregulator of COL7A1 in fibroblasts and keratinocytes, the regulation taking place primarily at the transcriptional level.
| Immunofluorescence | Anchoring Fibrils | |
|---|---|---|
| Skin | + | + |
| Chorioamnion | + | + |
| Placenta | − | − |
| Skeletal muscle | − | − |
| Cornea (Bowman membrane) | + | + |
| Oral mucosa | + | + |
| Cervix | + | + |
| Esophagus | + | + |
| Anal canal | + | + |
| Kidney cortex | − | − |
| Lung alvoli | − | − |
| Liver sinusoids | − | − |
| Stomach (fundus) | − | − |
| Large intestine | − | − |
| Elastic cartilage | − | − |
The biology of type VII collagen
Type VII collagen was initially described as an extended, unusually long molecule, hence the original designation as long-chain (LC) collagen. Rotary shadowing electron microscopy of type VII collagen molecules synthesized and secreted by human keratinocytes in culture revealed a long, 424-nm, triple-helical domain and flanking noncollagenous sequences ( Fig. 1 A). The amino-terminal domain was particularly noticeable, with individual α-chains contributing an extended arm. Simultaneously, visualization of type VII collagen isolated by limited pepsin proteolysis of amniotic membranes revealed a 780-nm dimer of 2 identical molecules in antiparallel orientation, with a 60-nm overlap stabilized by disulfide bonds ( Fig. 1 B, C). Further proteolytic digestion with pepsin revealed that type VII collagen molecules consist of a central collagenous, triple-helical segment flanked by the noncollagenous NC-1 and NC-2 domains. Subsequent cloning of the human type VII collagen gene and the corresponding complementary DNA (cDNA) indicated that the initially synthesized type VII collagen subunit polypeptide, the pro-α1(VII) chain is a complex modular protein consisting of a central, 1530-amino acid triple-helical domain ( Fig. 2 A). However, unlike interstitial collagens, the repeating Gly-X-Y sequence is interrupted by 19 imperfections because of insertions or deletions of amino acids in the Gly-X-Y repeat sequence. In the middle of the triple-helical domain, there is a 39-amino acid noncollagenous “hinge” region that is susceptible to proteolytic digestion with pepsin. The amino-terminal NC-1 domain of type VII collagen (NC-1[VII]), approximately 145 kDa in size, consists of submodules with homology to known adhesive proteins, including segments with homology to cartilage matrix protein, 9 consecutive fibronectin type III-like (FN-III) domains, a segment with homology to the A domain of von Willebrand factor, and a short cysteine and proline-rich region. The carboxy-terminal noncollagenous domain, NC-2, is fairly small, approximately 30 kDa, and it contains a segment with homology to the Kunitz protease inhibitor molecule (see Fig. 2 A).
Related posts:


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


