Skin Structure and Function
The integument comprises the skin together with its appendages ( Figs. 1.1 and 1.2 ). These include hair and hair follicles, sebaceous and sweat glands, and nails. The skin covers the entire body and is the largest organ of the body. It covers a surface area of more than 1.7 m 2 , making up in total about 16% of normal body weight. It has an array of functions. These include acting as a barrier to physical, biological and chemical agents, as well as to ultraviolet (UV) radiation. Skin barrier function also acts to control body hydration. Other functions include: sensory and thermoregulatory roles, vitamin D synthesis, immune surveillance, excretion of wastes through sweat glands, socio-sexual communication and reproduction, by virtue of its appearance and smell (e.g. hormones and pheromones).
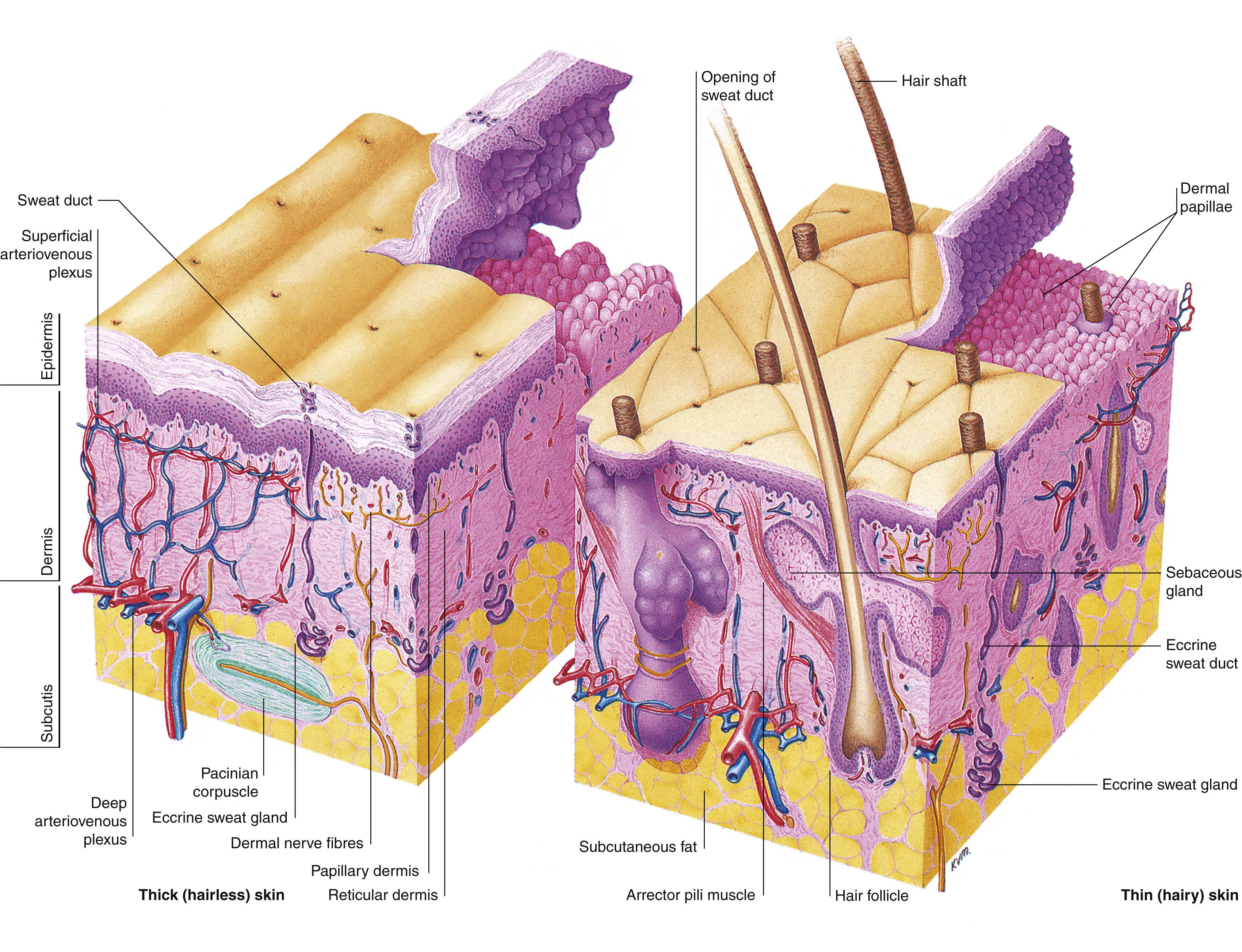
Skin is divided into glabrous (covering the palms of the hands and soles of the feet) and hairy skin. The skin comprises of two layers, the outermost epidermis and the innermost layer, the dermis. Embryologically, these two layers of skin are derived from the ectoderm and mesoderm, respectively. The epidermis and dermis are firmly attached to each other, and together vary in thickness from around 0.5 to 4 mm or more depending on body site. At the point where the epidermis meets the dermis, invaginations that project into the dermis are formed, known as “rete ridges” or “pegs.” Complementary projections of the dermis are called dermal papillae.
Epidermis
The epidermis is defined as a stratified squamous epithelium made up of several cell types. These include melanocytes (production of melanin pigment), Langerhans cells (immune function), Merkel cells (sensory function), and keratinocytes, of which the latter make up at least 80% of its cellular population.
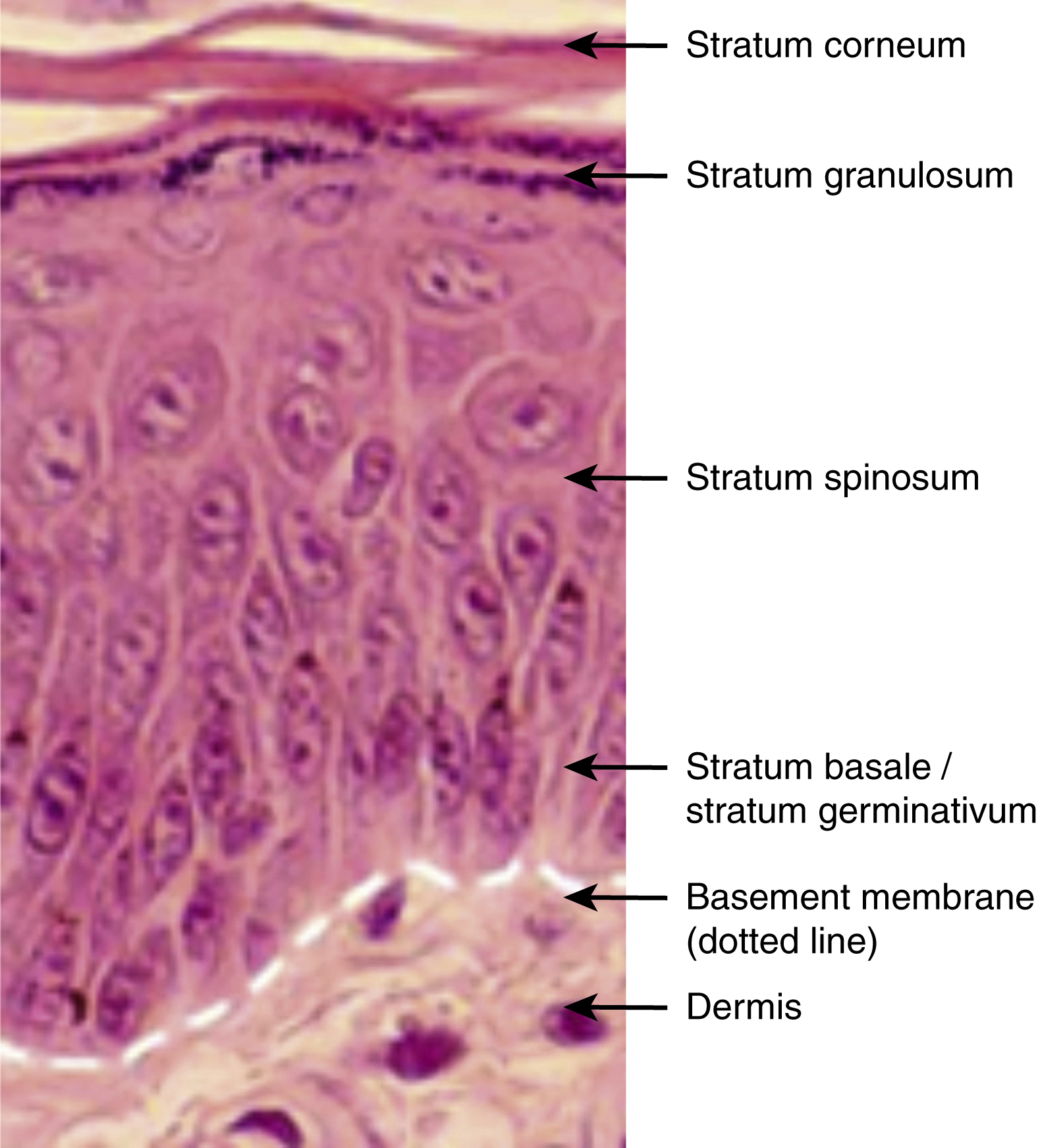
The epidermis is arranged into distinct layers, each showing a pattern of keratinocyte proliferation, differentiation, and maturation. Four main layers exist. These are strata basale (germinativum), spinosum, granulosum, and corneum. In thick skin, a fifth layer called the stratum lucidum is present and is found between the granular and the cornified layers. These layers reflect the sequential differentiation of keratinocytes as they migrate from the basal layer at the onset of terminal differentiation, having lost the ability to proliferate, to the outermost cornified layers where they are sloughed off. The process of terminal differentiation involves a series of biochemical and morphological changes, which result in the production of an anucleated cornified keratinocyte that forms the stratum corneum. Epidermal turnover takes on average 14–30 days.
The stratum basale is a single layer of cuboidal basal cells attached to the basement membrane by hemidesmosomes that contain integrins. Adjacent cells are attached by desmosomes that contain cadherins. The majority of the cells in this layer are mitotically active and are required for the continued renewal of the epidermis by upward displacement, replacing the cells of the outermost superficial layer that are lost during normal epidermal turnover. These mitotically active cells derive from a population of putative stem cells that are thought to reside in the deep rete ridges of glabrous skin and at the tips of the dermal papillae in interfollicular epidermis or in the bulge region of the outer root sheath (ORS) of adult human hair follicles in hairy skin. Also arising from stem cells in the basal layer are transit amplifying cells and post mitotic cells which are displaced into the suprabasal layers. Basal cells contain cytokeratins organized in bundles around the nucleus and insert into desmosomes peripherally.
Above this layer is the stratum spinosum, consisting of several layers of irregular, polyhedral shaped cells that display spiny projections. Cells from the previous layer lose contact with the basement membrane and are subsequently pushed up to form this one. Some cells in this layer are still mitotically active. Cells become progressively flattened as they move up towards succeeding layers. Cells contain lamellar granules (for the later provision of epidermal lipids responsible for the barrier properties of the stratified corneum) and more desmosomes for cell-to-cell adhesion.
The next layer, the stratum granulosum, comprises of three to five sheets of flattened cells. In this layer, as well as lamellar granules there are keratohyalin granules, which contain pro – filaggrin, a precursor of filaggrin that bundles the keratin filaments together. ,
The stratum lucidum consists of several layers of flattened cells without nuclei and organelles, and a keratin-rich cytoplasm.
The cornified layer consists of dead, anucleated, highly keratinized cells called squames or corneocytes. Keratin filaments polymerize by forming strong disulphide bonds. Filaggrin, a protein component of the keratohyalin granule, is involved in this process. The cornified envelope forms due to the catalytic activity of the enzyme transglutaminase which crosslinks proteins such as involucrin (an insoluble 70–80 kDa cysteine rich protein) in the plasma membrane. Other proteins found as components of the cornified cell envelope include keratolinin, loricrin, small proline-rich proteins, the serine proteinase inhibitor elafin, filaggrin linker-segment peptide, and envoplakin. Lipids, discharged by lamellar granules, fill the intercellular spaces and contribute to the barrier properties of the epidermis. The lack of desmosomes in the cells that are closest to the outermost layer results in the loss or shedding of corneocytes from the skin.
Keratins
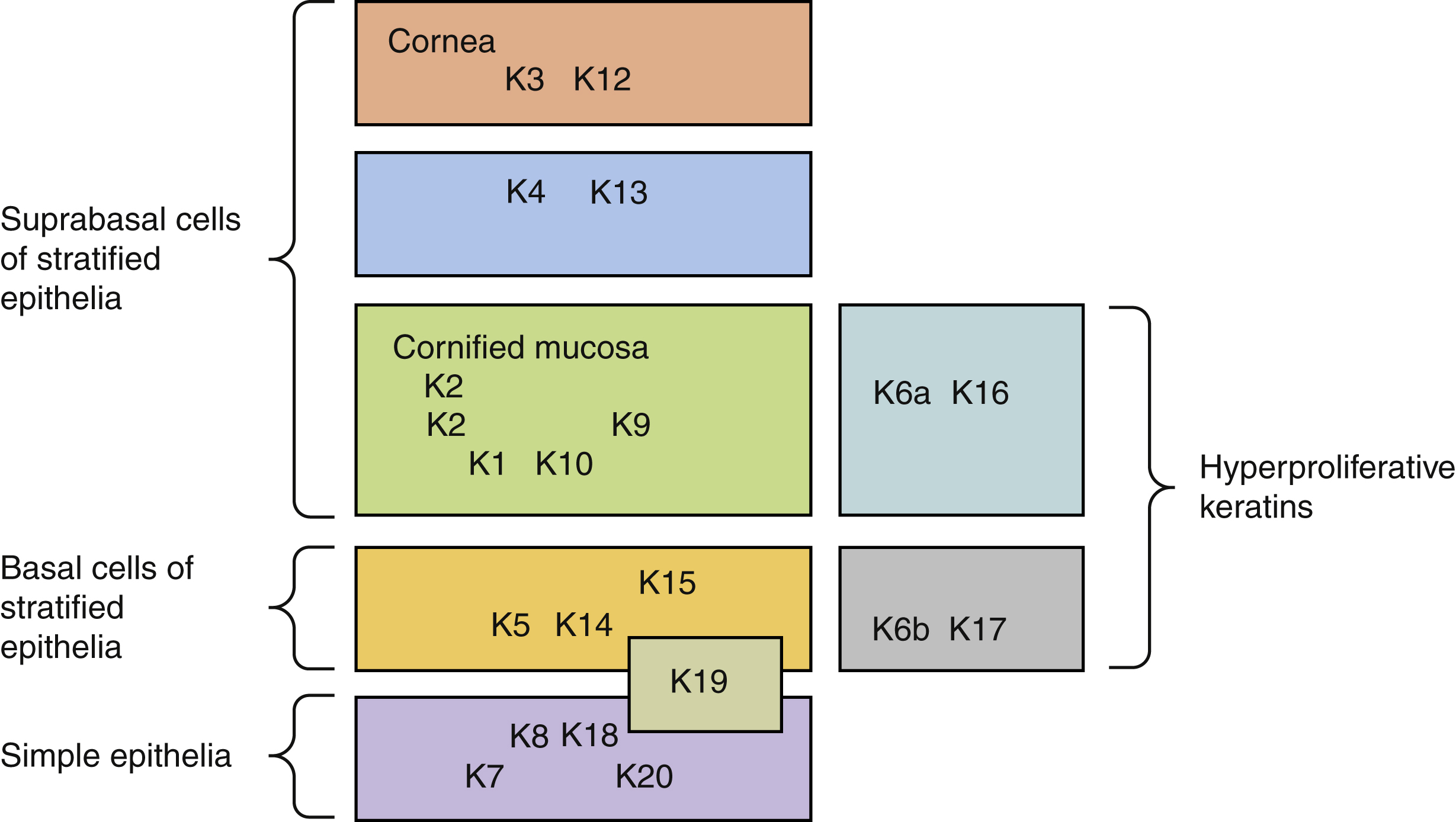
The cytoskeleton of all epithelial cells, including keratinocytes, is formed from the three groups of filaments, actin (microfilaments), tubulin (microtubules), and intermediate filaments. The keratins belong to the latter group, a multi-gene family of proteins that form filaments of 10 nm in diameter in which keratins form the two largest groups. These groups, which comprise more than 30 members of the keratin family, have been designated as type I keratins (acidic), numbered 9–20, and type II keratins (basic), numbered 1–8. Usually, type I and type II keratin subunits pair up and the heterodimers formed are expressed according to epithelial type and in a differentiation-state specific manner. For example, in simple epithelia, keratin K8 and K18 are expressed. Cells in the basal layer of stratified epithelia express K5 and K14. The differentiating suprabasal layers are characterized by the keratin pair K1 and K10. K6 and K16 are not found in normal epidermis except the outer root sheath of the hair follicle and junctional region. This keratin pair is also constitutively expressed in certain stratified squamous mucosal epithelia and the skin of the palm and sole. Expression of K6 and K16 is induced in wound healing epidermis, hyperproliferative epidermis such as psoriasis, squamous cell carcinomas, and hypertrophic scarring. Thus, keratin expression provides specific epithelial cell markers to assess epithelial proliferation and differentiation ( Fig. 1.3 ).
The Dermo-Epidermal Junction (Basement Membrane Zone)
The basement membrane zone lies at the interface between the epidermis and the dermis and strongly attaches these layers to each other. It also serves as a support for the avascular epidermis. Four layers have been identified within the basement membrane zone. These layers are:
- 1.
The plasma membrane of the basal keratinocyte, which is penetrated by hemidesmosomes.
- 2.
The electron-lucent lamina lucida that contains laminin, bullous pemphigoid glycoprotein, anchoring filaments and sub-basal dense plates.
- 3.
The electron-dense lamina densa that contains collagen type IV and proteoglycans.
- 4.
The collagen type VII rich-lamina fibroreticularis that contains anchoring fibrils ( Fig. 1.4 ).
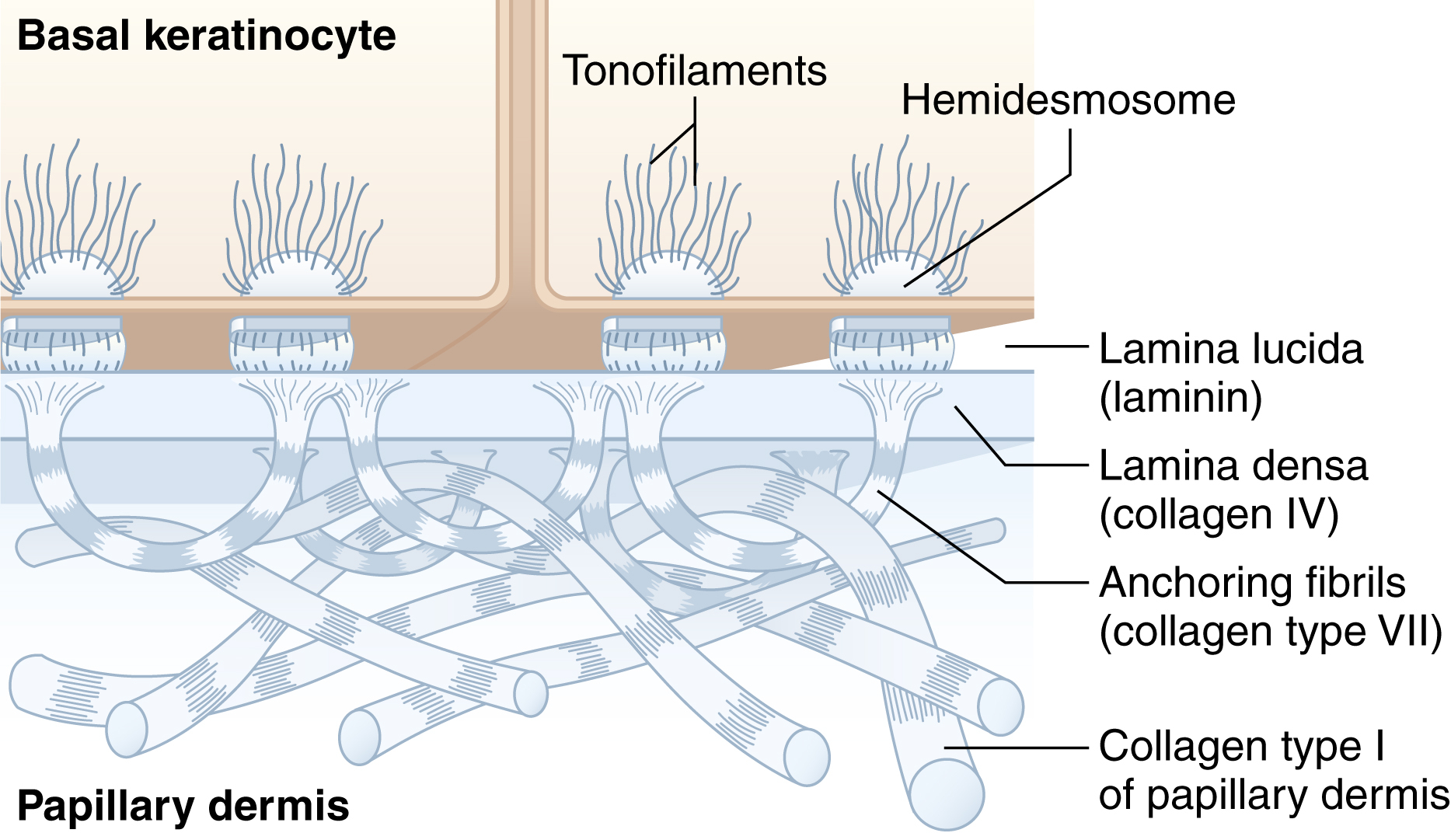
Fig. 1.4
The basement membrane zone.
Fibroblasts or keratinocytes are capable of synthesizing many of the components of the basement membrane zone. However, the combination of both cell types leads to the correct assembly of the complex.
Dermis
The dermis is thicker than the epidermis and is a dense, irregular fibroelastic connective tissue composed of extracellular matrix (ECM). The dermis is composed mostly of interwoven type I collagen with some type III collagen that provides the skin with its mechanical, tensile strength. It is subdivided into superficial papillary dermis that is in contact with the epidermis and deep reticular dermis. , The former is loosely woven and interdigitates with the epidermal ridges along the dermo-epidermal junction and contains thinner and more loosely arranged collagen and elastin fibers. It also contains more cells. The reticular dermis contains coarser and more tightly bound collagen fibers. At the arbitrary plane of this division there is a subpapillary plexus of blood vessels. Other ECM components making up the dermis include the amorphous ground substance of glycosaminoglycans (GAGs), namely, hyaluronic acid, dermatan sulphate, chondroitin-6-sulphate, chondroitin-4-sulphate, and heparin sulphate. Skin appendages pass through the dermis and may extend into the underlying subcutaneous fat. The dermis also has a complex lymphatic and nerve network. The resident cell types of the dermis include fibroblasts, macrophages, and mast cells.
Fibroblasts
The predominant cell type of the dermis is the fibroblast. These cells synthesize and degrade connective tissue matrix proteins (dermal remodeling) and a number of proteins that provide the structural ECM and basement membrane framework. These cells migrate on and between the surfaces of collagen fiber bundles. They also promote the interaction between the epidermis and dermis by the synthesis of soluble markers that are important in the regulation of normal skin physiology as well as during wound healing. Some of these growth factors/cytokines include keratinocyte growth factors (KGF) 1 and 2, transforming growth factor-β1 (TGF-β1), platelet-derived growth factor-A (PDGF-A), vascular endothelial growth factor (VEGF), and granulocyte/macrophage colony-stimulating factor (GM-CSF), allowing fibroblasts to engage in paracrine and autocrine interactions. Fibroblasts also exist as a highly diverse population. For example, the papillary dermis and reticular dermis have distinct fibroblast subpopulations with different characteristics.
The Extracellular Matrix
The extracellular matrix (ECM), also known as the connective tissue or mesenchyme, is composed of a mixture of proteins and polysaccharides in which cells are embedded. In the dermis, the matrix macromolecules are secreted largely by fibroblasts that also help to orientate the matrix. For example, fibroblasts can bind to collagen and exert force along a fiber, re-aligning it. Both the dermis and the basement membrane of skin represent diverse forms of ECM. The ECM functions to provide structural support and tensile strength, for cell adhesion and migration, and to regulate cell behaviour, influencing their development, migration, proliferation, shape and function. The ECM is composed of two main classes of extracellular macromolecules:
- 1.
Structural proteins and the adhesive proteins : among the structural proteins is collagen, the most abundant ECM component that provides tensile strength and helps to organize the matrix. Its deposition, degradation, and remodeling also appears to be tightly controlled. Elastin, another structural protein, functions to give resilience to the matrix. The adhesive proteins link components of the ECM both to one another and to the surfaces of cells. For example, laminin promotes the attachment of epithelial cells to the basement membrane, and fibronectin of fibroblasts and other cells to the ECM in the dermis.The major cell surface receptors responsible for the attachment of cells to the ECM are the transmembrane glycoproteins, integrins. The extracellular portion of integrins binds to collagens, laminins, and fibronectin; whereas the intracellular portion binds to the actin filaments of the cytoskeleton. This cytoskeleton-ECM linkage is responsible for the stability of cell–matrix junctions.
- 2.
Proteoglycans : the proteoglycans are polymorphic macromolecules consisting predominantly of molecules with a core protein and glycosaminoglycan (GAG) side chain extensions with repeating disaccharide units. They are divided into intracellular, membrane-bound, and extracellular groups. The extracellular molecules are relatively inflexible, taking up a spiral shape, and carry a negative charge, conferring hygroscopic and hydrostatic properties upon the ECM, and resist compressive forces. Proteoglycans regulate the function of several cellular processes, influencing cell proliferation and differentiation, and exert control over the behavior of the tissue as a whole under normal physiological conditions, during morphogenesis and during tissue repair processes. Some of these functions include organizing the ECM structure (GAGs and proteoglycans associate to form large complexes, for example aggregan; they can also associate with collagens and the basal lamina), ligand binding of potently active polypeptides modulating their bioactivity, cell adhesion, and transmembrane signaling.
Mesenchymal Stem Cells
The adult bone marrow contains mesenchymal stem cells (MSCs) (also known as marrow stromal cells) as well as hematopoietic stem cells (HSCs) (that generate all the blood cell lineages). MSCs are multipotent cells that have a high self-renewal capacity and give rise to many mesenchymal-derived tissues such as bone, cartilage, ligament, muscle tendon, adipose and marrow stroma. Human MSCs are fractionated from bone marrow aspirates by density centrifugation, collected from the density interface and cultured in the presence of fetal bovine serum specific for their expansion. The resulting cells in culture are a phenotypically homogeneous population. Human MSCs express surface antigens such as CD44, CD29, CD71 as analysed by flow cytometric analysis and are positive for the human MSC-specific antibodies such as SH-2 (for CD105) and SB-10 (for CD166). In vitro, these cells have been shown to undergo adipogenic, chondrogenic, and osteogenic differentiation depending on culture conditions.
Epithelial/Mesenchymal Interactions
Evidence is accumulating that the mesenchyme plays a crucial role in epidermal homeostasis. It is known that the epidermis, in turn, can influence the dermis with regards to structural remodeling, reinnervation, and vascularization. These epithelial/mesenchymal interactions are orchestrated by a complex network of communications between cell–matrix, cell–cell, and cell-diffusible factors.
The hair follicle provides a good system to study epithelial/mesenchymal interactions. For instance, the size and position of the developing hair follicles is determined by direct mesenchymal signals. Signaling between the dermal papilla and hair follicle epithelium is crucial for hair follicle development and function. The mesenchyme determines the fate of the epithelium. For example, the formation of epidermis and hair follicles is induced when cultured rabbit corneal epithelium is combined with embryonic hair-forming mouse dermis.
An understanding of the epithelial/mesenchymal interactions that takes place in skin arose from wound healing studies. Initially, Rheinwald and Green in 1975 showed the improved growth rates of keratinocytes grown on an irradiated mouse fibroblast feeder layer. Since then, the importance of a dermal component for the improved clinical take of cultured epithelium has been well documented. In vitro skin models have been used to study the importance of epithelial/mesenchymal interactions. Such organotypic cultures are composed of keratinocytes and mesenchymal cells grown in vitro on an extracellular matrix, and at an air–liquid interface to induce epidermal differentiation. They share similarities with wound healing, hyperproliferative and psoriatic skin, and have demonstrated the importance of epithelial/mesenchymal interactions in a number of processes including: epidermal homeostasis, production and deposition of basement membrane and ECM components, and secretion of diffusible factors.
Skin Aging
Aging occurs due to a combination of intrinsic and extrinsic causes. Intrinsic causes are due to less collagen and elastin being produced as a person ages, resulting in more fragile and thinner skin, thus forming wrinkles later in life. Dermatoporosis was first put forward as the name for chronic cutaneous fragility of the skin by Kaya and Saurat in 2007. With aging the epidermis thins as it develops rete ridges due to keratinocytes losing the ability to proliferate effectively and the epidermal turnover is slower. The dermis loses volume due to loss of hyaluronic acid (HA) in the extracellular matrix and the collagen is more disorganized, seeing an increase in the ratio of type 3 compared to type 1 collagen. Delayed wound healing is a prominent feature of aging skin. The reduced HA leaves the skin more vulnerable to tears following minimal trauma. Extrinsic factors include: sun (UV radiation) and environmental damage, such as tobacco smoking. Several studies have shown that cigarette smoke furthers wrinkle formation, nicotine also induces expression of matrix metalloproteinases which degrade the extracellular matrix in human skin. Severe sun damage causes epidermal atrophy, which is histopathologically distinguished by solar elastosis (degraded dermal elastic tissue).
Type 1 – Classical Cutaneous Wound Healing
A wound, in the context of skin, is a breach in the barrier that distinguishes an organism from its environment. The process through which the organism works in order to address this breach is “wound healing,” which, due to the important role the skin plays for the survival of the organism, is quite literally vital, and conserved through evolution. In the normal course of events, a lower species accepts tissue loss and heals a wound by exposure, licking, picking, and at the molecular level, scarring. The single most important impact on wound healing by man has been the early closure of wounds, by apposition with sutures in incisional wounds, and skin replacement in excisional wounds. Humans can deny significant skin and composite tissue loss by a “like for like” replacement in the specialty of plastic and reconstructive surgery, and here we can boast a form of “supranormal wound healing.”
Wound Healing Classifications
There are many ways to classify wound healing. In simple terms, we can consider four phases – coagulation, inflammation, fibroplasia, and remodeling; four types – fetal, adult, acute, and chronic; four ages – young, plateau, regressing, atrophic; and two systems of healing – epidermal and dermal. We can also classify wound healing in terms of clinical features and their wound management ( Table 1.1 ).
| Wound ± Intervention | Character Healing | |
|---|---|---|
| Type I: Classical | ||
| a) “Normal”/pimary incisional – incisional, no tissue loss | i) Closed early (<48 h) | Low risk of infection Minimal line scar |
| ii) Closed late (>48 h) | Increased risk infection Increased risk chronicity More significant scar | |
| iii) Unclosed | Healing by granulation (similar to Type Ibii), but in absence of tissue loss) High-risk chronicity | |
| b) “Normal”/secondary excisional – tangential tissue loss, no replacement | i) Above mid-dermis | Rapid re-epithelialization (<10 days) Minimal clinical scar |
| ii) Below mid-dermis | Slow/absent re-epithelialization High risk of infection and chronicity Closure by scar | |
| Type II: Neo-classical | ||
| a) “Supranormal” by skin replacement | i) Early split- or full-thickness skin grafting of Type Ibii | Present gold standards Cosmetic and functional problems remain Donor defect |
| ii) Biotechnological skin replacements (Cuono technique, in vitro composite grafts) | Ideally autologous and with viable cellular elements Opportunity to improve on Types Ibii and IIai | |
| b) “Supranormal” with apparent acceptance of tissue loss | i) Type Ibi treated with CKAllo or vapour permeable membrane | Follicle recruitment by activated extended follicle units |
| ii) Chronic full-thickness wound treated with CKAllo or vapour permeable membrane | Simulation of an acute wound environment | |
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


