FIGURE 7.1 Weak and strong μ-opioids were applied in the volar forearm of volunteers by dermal microdialysis. The intensity of opioid-induced maximum itch is shown in the left panel (visual analog scale: 0–10; mean ± SEM). Peak mast cell tryptase release during stimulation with the opioids is shown in the right panel (mean ± SEM). Only low-affinity opioids meperidine (40.4 mM) and morphine (3.11 mM) caused tryptase release from mast cells. The potent opioids alfentanil (1.2 mM), sufentanil (0.12 mM), and remifentanil (2.65 mM) provoked neither itch nor tryptase release. (Modified from Blunk, J. A, Schmelz, M., Zeck, S., Skov, P., Likar, R., and Koppert, W., Anesth Analg, 98, 364–370, 2004.)
Opioid Effects in Neurons and Keratinocytes
Opioid-induced itch has often been linked to peripheral release of histamine from mast cells as intradermally injected opioids can activate mast cells by a non-receptor-mediated mechanism (17). Accordingly, weak opioids, such as codeine, have been used as a positive control in skin prick tests. The opioid-induced release of histamine and mast cell tryptase can be specifically monitored by measuring tryptase concentration with dermal microdialysis following intraprobe delivery (18). In contrast to morphine, the highly potent μ-opioid agonist fentanyl does not provoke any mast cell degranulation, even if applied at concentrations having μ-agonistic effects exceeding those of morphine (Figure 7.1) (19). Thus, only high local concentrations of opioids are sufficient to degranulate mast cells and to induce itching regardless of their affinity to μ-opioid receptors. Therefore, local μ-opioid activation in the skin is not sufficient to provoke acute itch.
However, local opioid signaling in the skin has been identified as an important factor for epidermal homeostasis and including modulation of keratinocyte differentiation, skin barrier function, inflammation, and wound healing (20–22). Thus, therapeutic approaches using modulators of opioid signaling in the skin are not primarily directed toward neuronal effects but use these homeostatic interactions such as delta-opioids for improving skin barrier repair (23). Based on the close interactions between keratinocytes and sensory nerve endings in the epidermis, the cross talk between them (24–26) changes in keratinocyte differentiation, and mediator release based on changes in opioid signaling appears sufficient to cause sensitization and activation of the sensory nerves and thus underlie skin symptoms of itch and pain reported in sensitive skin.
Itch Induced by Nociceptors
Cowhage spicules inserted into the human skin produce itch in an intensity which is comparable to that following histamine application (27,28) but is not accompanied by an axon reflex erythema and is unresponsive to histamine (H1) blocker (29). The active compound cysteine protease muconain has been identified and has shown to activate proteinase-activated receptor 2 (PAR 2) and even more potently PAR 4 (10). Interestingly, mechanoresponsive polymodal C fiber afferents, the most common type of afferent C nociceptors in the human skin (30), can be activated by cowhage in the cat (31), in nonhuman primates (11,29), and in human volunteers (32) (Figure 7.2).
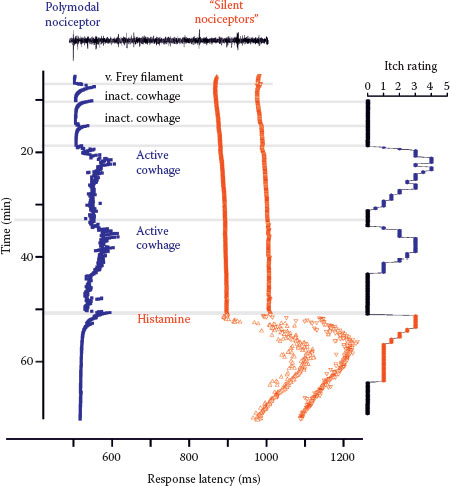
FIGURE 7.2 Specimen of a microneurography recording from the peroneal nerve in humans (raw signal with marked action potentials on top) showing action potential responses to electrical stimulation to successive electrical stimulation in the receptive field plotted from top to bottom. Mechanical stimuli (von Frey filament, inactivated cowhage spicules) only activate the polymodal nociceptor (blue squares) as seen by irregular increases of response latency, whereas the two mechano-insensitive silent nociceptors (orange-red symbols) are not activated. The polymodal nociceptor is activated by the application of active cowhage in parallel to the itch rating of the subject shown on the right panel. In contrast, the silent nociceptors do not respond to cowhage stimulation but are activated following histamine ionotophoresis. Itch ratings are given on a numerical rating scale from 0 (0 = no itch) to 10 (10 = maximal imaginable itch). Active cowhage and histamine evoke similar itch; however, the first appears to be encoded by polymodal nociceptors whereas the latter by histamine-sensitive silent nociceptors. (Modified from Tominaga, M., and Takamori, K., J Dermatol, 41, 205–212, 2014.)
Given that cowhage spicules can activate a large proportion of polymodal nociceptors, we face a major problem to explain why the activation of these fibers by heat or by scratching actually inhibits itch, whereas the activation by cowhage produces it.
Encoding Itch by Subpopulations of Nociceptors
Considering nociceptors being involved in generating itch, a population code has been postulated (pattern theory) (1,6,33) in which only a subpopulation of nociceptors can also be activated by pruritic stimuli, whereas pure nociceptors are only responsive to algogens. Accordingly, itch will be felt when only the first subpopulation is responding but pain will be felt when both populations are active.
The encoding of itch by nociceptors has also been proposed to rely on a spatial code (34) based on the itch induced by capsaicin being locally applied on a cowhage spicule into the epidermis (28). The highly localized stimulation in the epidermis strongly activates some of the local nociceptors, while their immediate neighbors remain silent, resulting in a mismatch signal of activation and absence of activation from this site. It has thus been hypothesized that this mismatch might be perceived by the CNS as itch (32,34). Therefore, it needs to be pointed out that pruritus cannot only be explained by itch-specific or itch-selective neurons (7) along the specificity theory. In addition, the pure spatial pattern of activated nociceptors might similarly underlie the itch sensation without any requirement of itch-specific primary afferent neurons.
Peripheral Sensitization in Itch and Pain
Spontaneous itch and pain are of paramount clinical relevance, as they correlate with the main complaint of patients with chronic itch and pain. It is highly interesting that the patterns of sensitization linked to chronic pain and itch are remarkably similar.
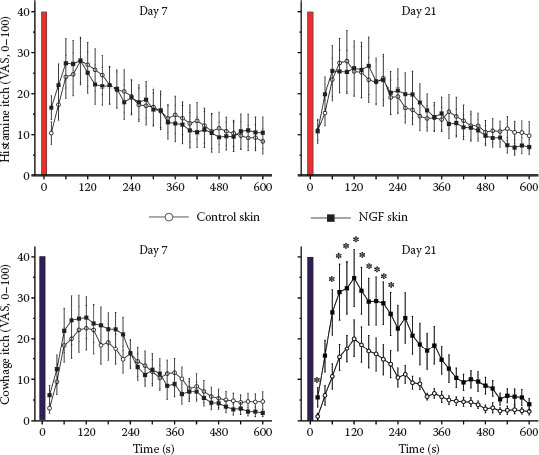
FIGURE 7.3 Itch ratings of human volunteers following stimulation with histamine (upper panels) and cowhage spicules (lower panels) are shown. The stimulations were applied in the forearm skin injected with saline (control) or NGF (1 μg in 50 μL) 7 days (left panels) or 21 days (right panels) after the injections. NGF injections did not sensitize histamine-induced itch, but cowhage-induced itch was increased by NGF at the maximum of the mechanical hyperalgesia (21 days; right lower panel). (Modified from Rukwied, R. R., Main, M., Weinkauf, B., and Schmelz, M., J Invest Dermatol, 133, 268–270, 2013.)
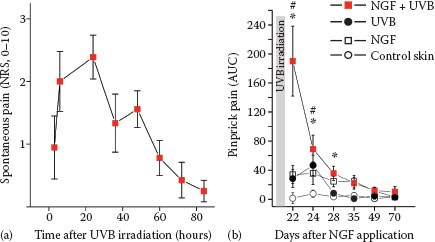
FIGURE 7.4 (a) Presensitization with NGF (1 μg) injected 3 weeks before UVB irradiation (threefold minimum erythema dose) provoked spontaneous pain in subjects with a time course matching the intensity of the UV-induced inflammation. (b) Hyperalgesia to pinprick stimuli develops following intradermal NGF injection and also for about 3 days after UVB irradiation. Combined sensitization with NGF and UVB irradiation causes a supraadditive increase of mechanical hyperalgesia. (Reprinted from Biochimie, 92, Okudaira, S., Yukiura, H., and Aoki, J., Biological roles of lysophosphatidic acid signaling through its production by autotaxin, 698–706, Copyright (2010), with permission from Elsevier.)
There is cumulative evidence for a prominent role of NGF-induced sensitization of primary afferents in both chronic itch and pain; increased levels of NGF were found in patients with chronic itch suffering from atopic dermatitis or psoriasis (35–38). Similarly, there is clear evidence for a major role of NGF in chronic inflammatory pain (39–41). Moreover, blocking NGF by specific antibodies proved to be analgesic in the patients with chronic pain (42,43). Anti-NGF strategies were also successful in animal models of chronic itch (44). It is therefore not surprising that intradermally injected NGF not only causes hyperalgesia to heat and mechanical stimuli in volunteers (45,46), but also sensitizes for cowhage-induced itch (47) (Figure 7.3).
Intracutaneous NGF injection does not induce visual inflammatory responses in humans (46), but interestingly, when combined with an inflammatory pain model (ultraviolet B [UVB] sunburn), the subjects report spontaneous pain (Figure 7.4) and pronounced hyperalgesia (48) that also includes axonal hyperexcitability (49). These results nicely match the analgesic effects of anti-NGF in chronic inflammatory pain that are not accompanied by reduced signs of inflammation (42). Therefore, it emerges that neurotrophic factors such as NGF can change expression patterns of primary afferent nociceptors such that their ability to signal pain or itch by local inflammatory mediators is increased. This increase might be based on higher discharge frequencies linked not only to sensitized transduction, but also to axonal hyperexcitability.
Links between Sensitization Mechanisms in Chronic Itch and Sensitive Skin
The structural damage of neurons can lead to hyperexcitability resulting in neuropathic itch and pain. The neuropathy of skin nerves can be quantified functionally by quantitative sensory testing or structurally by staining nerve fibers in skin biopsies (50). The correlation between structural loss of skin nerve fibers and reduced sensory function is well established (51). Unexpectedly, the degree of neuropathy does not predict the level of pain or itch symptoms (52,53). Functional and structural impairments are therefore essential to diagnose neuropathy (54). However, in most cases, neuropathy will simply lead to diminished sensory function without spontaneous pain or itch, for example, in diabetic neuropathy. Among patients with neuropathy, those with pain do not separate from those without pain based on the pattern or the degree of their neuropathy (52,55,56).
TABLE 7.1
Characteristics of Neuropathic and Inflammatory Pain as Compared to Sensitive Skin