Introduction
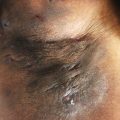
Hidradenitis suppurativa (HS) is an inflammatory condition characterized by painful and sometimes purulent nodules, abscesses, and sinus tracts located in intertriginous regions. Lesions may progress to form hypertrophic scars or dermal tunnels. HS patients are regarded as having one of the worst qualities of life among the major dermatologic conditions. Comorbidities include obesity, asthma, acne, diabetes, dyslipidemia, hypertension, thyroid disease, rheumatoid arthritis, depression, psoriasis, and polycystic ovarian syndrome.
The hypothesized pathogenesis of HS is multifaceted. On a cellular level, it is characterized by follicular hyperkeratosis which leads to a deleterious pathway involving follicular occlusion, dilation, inflammation, and rupture. The etiology involves a complex interplay between a dysregulated immune system and genetic, environmental, and microbiological factors.
Given the morbid nature of this disease, its etiology is of great interest to both patients and clinicians alike. The question “to what extent is HS caused by genetic factors?” was recently identified as one of the top 10 most important uncertainties in a Priority Setting Partnership, in which patients and clinicians agreed on mutually important HS research questions. While poor quality of life greatly affects these patients, they are more often worried about passing this condition on to their offspring.
Many features of HS support a genetic basis. Thirty to 42% of HS patients report a family history of HS. Family history of HS is significantly higher in early onset HS, which is also associated with more widespread involvement. There is an increased incidence of HS in monogenic disorders such as Dowling-Degos disease, Down syndrome, and keratitis, ichthyosis, and deafness syndrome, among others. Different HS phenotypes (axillary–submammary, follicular, and gluteal) show preferences in regard to sex predominance, age of onset, lesion type, location affected, and disease severity suggesting genetic heterogeneity. 17(p53) HS is also associated with many inflammatory diseases that have known polygenic etiologies, such as inflammatory bowel disease.
This chapter provides a general overview of the HS genetics and epigenetics literature. First, a review of HS investigational genetic studies is presented, describing mutations reported to be associated with HS susceptibility, phenotype, or treatment response ( Table 12.1 ). These studies are discussed according to study design: family linkage analyses, genome-wide association studies (GWAS), or target candidate gene studies. Genetic findings in syndromic HS—HS presenting as part of or along with a medical syndrome—are reviewed as well ( Table 12.2 ). Early work on HS epigenetics and microRNAs is discussed next. The chapter concludes by discussing the biological and clinical relevance of these findings as well as areas for future study.
| Study | Gene | Protein | Function | Identified Mutation(s) | Ethnicity | Familial/sporadic |
|---|---|---|---|---|---|---|
| Genome-Wide Linkage Analysis/Unbiased Linkage Analysis Studies | ||||||
| Gao et al., 2006 | Susceptibility locus 1p211–1q25.3 | Han Chinese | Familial | |||
| Wang et al., 2010 | NCSTN PSENEN PSEN1 | Nicastrin Presenelin enhancer 2 Presenelin 1 | Cofactor subunit of γ-secretase, a glycoprotein transmembrane aspartyl protease cleaving over 30 type I integral membrane proteins, including Notch and Amyloid Precursor Protein Cofactor subunit of γ-secretase Catalytic subunit of γ-secretase | 1 splice site, 1 truncating, 1 nonsense 1 truncating, 1 frameshift 1 truncating | Han Chinese | Familial |
| Liu et al., 2011 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 splice site, 1 truncating | Han Chinese | Familial |
| Chen et al., 2014 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 nonsense | African American | Familial |
| Faraji Zonooz et al. 2016 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 nonsense | Iranian | Familial |
| Takeichi et al., 2020 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 missense | Japanese | Familial |
| Genome-Wide Association Study | ||||||
| Liu et al., 2019 | BCL2 | BCL-2 | Anti-apoptotic regulatory protein involved in skin homeostasis | 5 single nucleotide polymorphisms associated with a positive response of HS to adalimumab | Not reported | Not reported |
| Target Candidate Gene Studies | ||||||
| NCSTN Mutations | ||||||
| Li et al., 2011 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 nonsense, 1 missense | Chinese | Familial |
| Pink et al., 2011 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 splice site | British | 42% familial |
| Pink et al., 2012 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 missense, 2 splice site | Not reported | Sporadic |
| Haines et al., 2012 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating | Singapore | Sporadic |
| Miskinyte et al., 2012 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 splice site, 2 truncating | French | Familial |
| Zhang et al., 2013 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 2 missense | Chinese | Familial |
| Nomura et al., 2013 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 splice site | Japanese | Familial |
| Jiao et al., 2013 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 missense | Han Chinese | Familial |
| Ma et al., 2013 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating | Chinese | Familial |
| Nomura et al., 2014 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating | Japanese | Familial |
| Yang et al., 2015 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating | Chinese | Familial |
| Ratnamala et al., 2016 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating | Indian | Familial |
| Zhang et al., 2016 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 missense | Chinese | Familial |
| Liu et al., 2016 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating, 1 missense | Caucasian | Familial, Nonfamilial |
| Xiao et al., 2018 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 frameshift | Sporadic | Familial |
| Wu et al., 2018 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 truncating | Chinese | Familial |
| PSENEN | ||||||
| Pink et al., 2011 | PSENEN | Presenelin enhancer 2 | Cofactor subunit of γ-secretase | 1 frameshift | British | Familial |
| Zhou et al., 2016 9(p20) | PSENEN | Presenelin enhancer 2 | Cofactor subunit of γ-secretase | 1 missense, 1 splice site | Chinese | Familial |
| Kan et al., 2018 | PSENEN | Presenelin enhancer 2 | Cofactor subunit of γ-secretase | 1 frameshift | Japanese | Familial |
| Mutations in Other Genes | ||||||
| Savva et al., 2013 | TNF | TNF-α | Pro-inflammatory cytokine elevated in HS lesional skin | SNP (-238G/A) in promoter region linked to HS susceptibility | Not reported | 29% familial |
| Giatrakos et al., 2013 | IL12Rβ1 | IL12Rββ1 receptor subunit | Part of IL-12 receptor complex; IL-12/IL-23 pathway involved in antigen presentation in HS | Haplotype (h2) associated with more severe disease vs. haplotype (h1) associated with an older age of onset | Caucasian | Sporadic |
| Giamarellos-Bourboulis et al., 2016 | DEFB (cluster of nine β—defensing genes) | β-defensin | Proinflammatory mediators; deposits of β-defensin-2 and -3 found in HS lesional skin | > 6 copy number of DEFB (cluster of 9 beta-defensin genes) associated with a 6.72 OR for HS ( P < .0001) | Greek (cohort 1), German (cohort 2) | 20.9% and 37.1% familial in cohort one and two, respectively |
| Agut-Busquet et al., 2018 | MYD88 | Myeloid differentiation primary response protein | Involved in innate and adaptive immune response and Toll-like receptor and IL-1 receptor signaling | SNP (GG genotype of rs6853) associated with an increased risk of severe HS | Caucasian | Not reported |
| Study | Gene | Protein | Function | Identified Mutation(s) | Ethnicity | Familial/Sporadic |
|---|---|---|---|---|---|---|
| HS + Nevus Comedonicus Syndrome (NCS) | ||||||
| Higgins et al., 2017 | FGFR2 | Fibroblast growth factor receptor 2 | Expressed in keratinocytes, hair follicles, and sebaceous glands; substrate-receptor binding results in cell division and differentiation | 1 missense | Unknown | Unknown |
| HS + Familial Mediterranean Fever | ||||||
| Vural et al., 2019 | MEFV | Pyrin | Involved in regulating inflammasome activity | Frequency of pathogenic MEFV variants associated with risk of severe and complex HS | Not reported | Not reported |
| HS + Darier’s Disease | ||||||
| Ornelas et al., 2016 | ATP2A2 | Sarcoplasmic reticulum calcium adenosine triphosphatase (SERCA) transport ATPase type 2 isoform pump | Important in maintaining calcium homeostasis; SERCA inhibition may affect Notch signaling | ATP2A2 mutation | Not reported | Not reported |
| HS + Dent Disease 2 | ||||||
| Marzuillo et al., 2018 | OCRLI | Inositol polyphosphate 5-phosphatase (OCRL1) | Forms a complex at maturing epidermal-dermal junction and regulates phosphoinositol-4,5-bisphosphate abundance (levels linked to inflammation) | OCRLI mutation | Not reported | Not reported |
| HS + Pachyonychia Congenita | ||||||
| Pedraz et al., 2008 | KRT6A | Keratin 6A | Provides strength to skin, nails, and hair | 1 missense | Not reported | Not reported |
| HS + Down Syndrome | ||||||
| Borbujo Martínez et al., 1992, Mengesha et al., 1999, Mebazaa et al., 2009, Blok et al., 2016 | Chr 21 | Primarily encodes Amyloid Precursor Protein | Integral membrane protein; competitive substrate for γ-secretase along with Notch | Trisomy 21 | Not reported | Sporadic |
| HS + Keratitis-Ichthyosis-Deafness (KID) | ||||||
| Montgomery et al., 2004, Lazic et al., 2012 | GJB2 gene, G12R gene | Connexin-26 protein | Forms gap junctions key for tissue homeostasis, growth, and development | 2 missense | Not reported | Sporadic |
| HS + Dowling-Degos Disease (DDD) | ||||||
| Zhou et al., 2016 | PSENEN | Presenelin enhancer 2 | Cofactor subunit of γ-secretase | 1 missense, 1 splice site | Chinese | Familial |
| Ralser et al., 2017 | PSENEN | Presenelin enhancer 2 | Cofactor subunit of γ-secretase | 2 splice site, 2 nonsense | 3 German, 2 Indian, 1 French patient | Not reported |
| Pavlovsky et al., 2018 | PSENEN | Presenelin enhancer 2 | Cofactor subunit of γ-secretase | 1 missense | Ashkenazi Jewish | Familial |
| González-Villanueva et al., 2018 | POFUT1 | Protein O-Fucosyltransferase 1 | Protein in endoplasmic reticulum; adds sugar moieties to Notch receptors | 1 splice site | Not reported | Not reported |
| PASH (Pyoderma Gangrenosum, Acne, and Suppurativa Hidradenitis) and PAPASH Syndrome | ||||||
| Braun-Falco et al., 2012, Marzano et al., 2013, 42(p201) Calderón-Castrat et al., 2016, Saito et al., 2018 | PSTPIP1 | Proline-serine-threonine phosphatase-interacting protein 1 | Involved in regulating inflammasome activity | 3 missense and 1 SNP with increased repetition of CCTG micro-satellite motif | Patient 1 Russian, Patient 2 German | Sporadic |
| Marzano et al., 2013 | NLRP3 IL1RN MEFV NOD2 PSMB8 | Cryopyrin IL-1 receptor antagonist Pyrin Nucleotide-binding oligomerization domain-containing protein 2 Proteasome 20S Subunit Beta 8 | Initiates and regulates the immune system’s response to injury, invasion, or toxins Modulates the IL-1 mediated immune responses Regulates inflammasome activity Involved in autophagy; when triggered by a bacterial antigen, it activates NFkB that regulates the immune response Subunit of immunoproteosome important in detecting self v. non-self proteins in immune cells to guide immune response | 1 missense 1 missense 2 missense 2 missense 1 missense | Not reported | Not reported |
| Duchatelet et al., 2015 | NCSTN | Nicastrin | Cofactor subunit of γ-secretase | 1 frameshift | Not reported | Two familial and one sporadic |
| Follicular Occlusion Triad | ||||||
| Musumeci et al., 2019 | KR17 | Keratin 17 | Provides strength to skin, nails, and hair | 1 missense | Not reported | Familial |
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


