19 The colour of normal skin comes from a mixture of pigments. Untanned Caucasoid skin is pink, tinted from white by oxyhaemoglobin in the blood within the dermis. Melanin (see Melanogenesis) blends with this colour, and may be increased, for example, after a suntan. Melanin is, of course, also responsible for the shades of brown seen not only in Congoid (Negroid) skin, but also in the other races. Various hues are caused by the addition to these pigments of yellow from carotene, found mainly in subcutaneous fat and in the horny layer of the epidermis. There is no natural blue pigment; when blue is seen, it is either because of an optical effect from normal pigment (usually melanin) in the dermis, or the presence of an abnormal pigment. Skin pigmentation (measured by skin reflectance) is darkest near the equator and correlates with latitude and ultraviolet radiation (UVR). Skin colour seems to have evolved as a compromise between being dark enough to block the damage to DNA caused by UVR and photolysis of the essential metabolite, folate, and light enough to allow vitamin D to be synthesized in the skin (p. 12). Melanins protect against UVR damage by absorbing and scattering the rays, and by scavenging free radicals. Hair colour is determined by the relative amounts of the different types of melanin (see Melanogenesis). Eumelanin predominates in black hair and phaeomelanin in red. Melanin is formed from the essential amino acid phenylalanine through a series of enzymatic steps in the liver and skin. Tyrosine, formed in the liver by hydroxylation of the essential amino acid phenylalanine under the influence of phenylalanine hydroxylase, is the substrate for the reactions that occur in melanocytes (Figure 19.1). Melanocytes are the only cells in the epidermis to contain tyrosinase (dopa oxidase), the rate-limiting enzyme in melanogenesis. Phaeomelanins and trichochromes, the pigments in red hair, are synthesized in a similar way, except that cysteine reacts with dopaquinone and is incorporated into the subsequent polymers. Phaeomelanins and eumelanins may intermesh to form mixed melanin polymers. Figure 19.1 The control of melanogenesis. Melanocortin 1 receptor (MC1R) activity is both constitutive and rate limiting when promoting melanogenesis, via cyclic adenosine monophosphate (cAMP) production and tyrosinase stimulation. The MC1R is activated by ligands such as α-melanocyte-stimulating hormone (α-MSH) and other pituitary peptides. In the absence of such ligands or the MC1R itself (knockout animals), and with loss-of-function mutations of the MC1R, phaeomelanin is produced. The precise mechanism by which ultraviolet radiation stimulates melanogenesis remains uncertain. Eumelanins and phaeomelanins differ from neuromelanins, the pigments found in the substantia nigra and in cells of the chromaffin system (e.g. adrenal medulla, sympathetic ganglia). The latter are derived from tyrosine using a different enzyme, tyrosine hydroxylase, which is not found in melanocytes. Melanin is made within melanosomes (see Figure 2.6), tiny particles measuring about 0.1 × 0.7 μm, shaped either like American footballs (eumelanosomes, containing eumelanin) or British soccer balls (phaeomelanosomes, containing phaeomelanin). Eventually, fully melanized melanosomes pass into the dendritic processes of the melanocyte to be injected into neighbouring keratinocytes. Once there, the melanosomes are engulfed in lysosomal packages (melanosome complexes) and distributed throughout the cytoplasm. Such secretory lysosomes are common to various haematopoietic cells and melanocytes. This explains why some genetic disorders of pigmentation (e.g. rare forms of albinism such as the Hermansky–Pudlak and Chediak–Higashi syndromes, p. 271) are linked with abnormal immune function. All of us, regardless of race or skin colour, have similar density of melanocytes, with one melanocyte supplying melanin to about 30 keratinocyte neighbours. What determine a person’s skin colour is the activity of those melanocytes and their interactions with their keratinocyte neighbours. Darker skin individuals have melanocytes that produce more and larger melanosomes. In addition, these melanosomes are more efficiently transferred to keratinocytes and more slowly degraded in the melanosome complexes. Melanogenesis can be increased by several stimuli, the most important of which is UVR. Tanning represents a protective mechanism by our skin against future UV damage and involves two distinct reactions. A neat control mechanism involving glutathione has been postulated. Reduced glutathione in the epidermis, produced by the action of glutathione reductase on glutathione, inhibits tyrosinase. UVR and some inflammatory skin conditions may induce pigmentation by oxidizing glutathione and so blocking its inhibition of melanogenesis. Melanocytes are also influenced by MSH peptides from the pituitary and other areas of the brain (Figure 19.1). However, these MSH peptides may play little part in the physiological control of pigmentation. Hypophysectomy will not cause a black skin to lighten and only large doses of adrenocorticotrophic hormone (ACTH), in pathological states (p. 276), will increase skin pigmentation. In the skin, α-MSH also acts as an anti-inflammatory agent by antagonizing the effects of interleukin 1 (IL-1) in inducing IL-2 receptors on lymphocytes (p. 18) and in inducing pyrexia. Oestrogens and progestogens (and possibly testosterone too) may, in some circumstances, stimulate melanogenesis, either directly (by acting on oestrogen and progestogen receptors in the melanocyte) or by increasing the release of MSH peptides from the pituitary. Genetic differences determine the pigmentation of the different races (see Chapter 14, p. 197). A black person living in Britain, and a white person living in Africa will remain black and white, respectively. None the less, there is some phenotypic variation in skin colour (e.g. tanning after sun exposure). Red hair is the result of genetic variations in the amino acid sequence of the melanocortin 1 receptor (MC1R; Figure 19.1). Some genodermatoses with abnormal pigmentation are described in Chapter 24. These may be caused by an imbalance of the normal pigments mentioned above (e.g. in cyanosis, chloasma and carotenaemia) or by the presence of abnormal pigments (Table 19.1). Sometimes, it is difficult to distinguish between the colours of these pigments (e.g. the gingery brown colour of haemosiderin is readily confused with melanin). Histological stains may be needed to settle the issue. In practice though, apart from tattoos, most pigmentary problems are caused by too much or too little melanin. Table 19.1 Some abnormal pigments. Some conditions in which there is a lack of melanin are listed in Table 19.2. A few of the more important, and the mechanisms involved, are summarized in Figure 19.2. Decreased melanin pigmentation can be caused by the absence of melanocytes (vitiligo) or abnormalities in melanin synthesis (albinism). It is sometimes difficult to distinguish between hypomelanosis (decreased melanin) and amelanosis (complete absence of melanin). Wood’s lamp (UVA light) examination can help enhance the contrast; the greater the loss of epidermal pigment, the more marked the contrast. Table 19.2 Some causes of hypopigmentation. Figure 19.2 The mechanisms involved in some types of hypopigmentation. Various genetic conditions exist in which there is a defect in the synthesis or packaging of melanin in the melanocyte, or a defective transfer of melanosomes to surrounding keratinocytes (see Chapter 2). In the most common type, little or no melanin is made in the skin and eyes (oculocutaneous albinism) or in the eyes alone (ocular albinism, not discussed further here). The prevalence of albinism of all types ranges from 1 in 20 000 in the United States and United Kingdom to 5% in some communities. The hair bulb test (see Investigations) separates oculocutaneous albinism into two main types: tyrosinase-negative and tyrosinase-positive. Roughly equal numbers of the two types are found in most communities, both being inherited as autosomal recessive traits. This explains how children with two albino parents can sometimes themselves be normally pigmented, the genes being complementary in the double heterozygote (Figure 19.3).
Disorders of Pigmentation
Normal skin colour
Melanogenesis
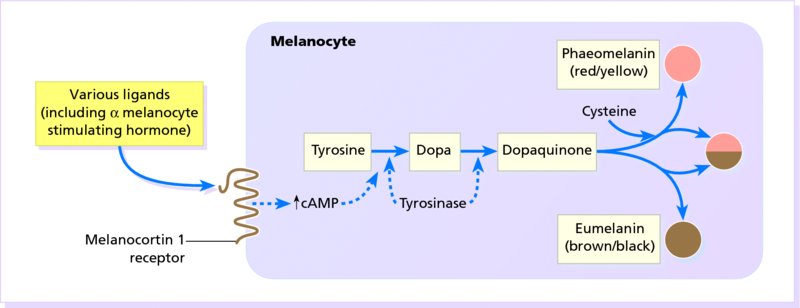
The control of melanogenesis
Genetics and skin pigmentation
Abnormal skin colours
Endogenous
Haemoglobin-derived
Methaemoglobin
Blue colour in vessels
Sulphaemoglobin
Cyanosis
Carboxyhaemoglobin
Pink
Bilirubin
Yellow–green
Biliverdin
Haemosiderin
Brown
Drugs
Gold
Blue–grey (chrysiasis)
Silver
Blue–grey (argyria)
Bismuth
Grey
Mepacrine
Yellow
Clofazamine
Red
Phenothiazines
Slate-grey
Amiodarone
Blue-grey
Diet
Carotene
Orange
Exogenous
Tattoo pigments
Carbon
Blue–black
Coal dust
Blue–black
Cobalt
Blue
Chrome
Green
Cadmium
Yellow
Mercury
Red
Iron
Brown
Local medications
Silver nitrate
Black
Magenta paint
Magenta
Gentian violet
Violet
Eosin
Pink
Potassium permanganate
Brown
Dithranol (anthralin)
Purple
Tar
Brown
Iodine
Yellow
Decreased melanin pigmentation
Genetic
Albinism
Piebaldism
Phenylketonuria
Waardenburg’s syndrome
Chediak–Higashi syndrome: autosomal recessive lysosomal defect, pale skin with sparse silvery-grey or blond hair, susceptible to infections
Tuberous sclerosis (p. 345)
Endocrine
Hypopituitarism
Chemical
Contact with substituted phenols (in rubber industry)
Chloroquine and hydroxychloroquine
Post-inflammatory
Eczema
Pityriasis alba
Psoriasis
Sarcoidosis
Lupus erythematosus
Lichen sclerosus et atrophicus
Cryotherapy
Infections
Leprosy
Pityriasis versicolor
Syphilis, yaws and pinta
Tumours
Halo naevus
Malignant melanoma
Miscellaneous
Vitiligo
Idiopathic guttate hypomelanosis 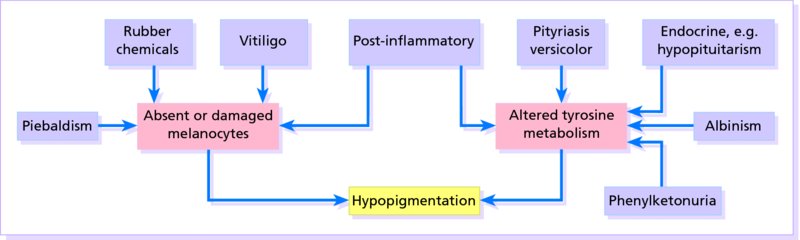
Oculocutaneous albinism
Cause
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


