CHAPTER 47 Harnessing Stem Cell–Mediated Lymphangiogenesis for the Treatment of Lymphedema
KEY POINTS
New molecular understanding of the growth and development of the lymphatic system is creating new opportunities for molecular intervention in lymphangiogenesis.
Lymphatic growth in tumors is one area of research that is revealing new insights into lymphatic growth and development, including the fact that tumor-associated macrophages can become lymphatic endothelial cells.
Tissue engineering approaches to the restoration of collecting lymphatics may hold promise for the treatment of lymphedema.
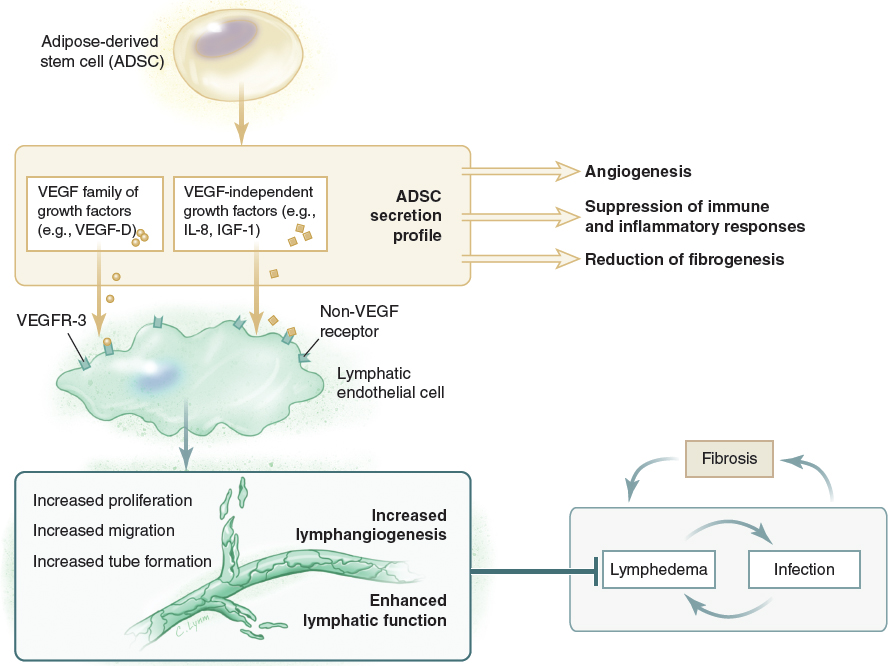
Adipose-derived mesenchymal stem cells appear promising for the treatment of lymphedema. However, their effectiveness, if confirmed, is probably the result of their ability to release growth factors in a sustained manner rather than their ability to differentiate into lymphatic tissue components.
Recent advances in the molecular biology, visualization techniques, and genetic modeling of the lymphatics have shed light on the developmental origins of this intricate vascular system. 1 , 2 This in turn has created new opportunities for the manipulation of lymphatic biology for clinical benefit, including the exploitation of protein growth factors, small molecules, and neutralizing antibodies to target the lymphatic vasculature. 3 In 2002 Niklason et al 4 stated that tissue engineering approaches for lymphatic engineering were “currently nonexistent.” Since then, several approaches, including the use of lymphangiogenic growth factors, lymphatic grafts, tissue transfers, and mesenchymal stem cells (MSCs), have largely been used experimentally and occasionally clinically to regenerate lymphatic vessels and their networks for the treatment of secondary lymphedema. The engineering of new tissues, including new lymphatics, requires a combination of cells, matrix, and growth factors. MSCs and in particular adipose-derived stem cells (ADSCs) are a promising source of cells for lymphatic regeneration, although more recent evidence indicates that their real value may be in their ability to sustainably release a range of growth factors. 5
Cellular Origin of Lymphatic Endothelial Cells
Lymphatic endothelial cells (LECs) are the most important cell type in lymphatic regeneration and therefore the key to any restoration of lymphatic function in lymphedema. The earliest LECs in embryonic development arise from primitive venous endothelial cell precursors 2 , 6 in the anterior cardinal veins and a separate superficial venous plexus. 7 – 9 The initial sprouting of the primitive lymphatic vessels is preempted by a transition from venous to lymphatic endothelial expression profiles of distinct LEC markers. 9 In murine models, Sry-related HMG box containing-18 (SOX18) and chicken ovalbumin upstream promoter or NR2F2 (COUP-TFII) expressing cells cluster into a prelymphatic subsection of the veins from 14.5 days to postcoitus. 7 The resulting jugulodigastric lymphatic sacs then give rise to collecting lymphatics, whereas the more peripherally derived LECs, which form the more superficial subtypes of lymphatics, continue to expand and only mature during later developmental stages in some areas postpartum. 3 , 8 Therefore endothelial cells from the initial and collecting lymphatics have a distinct origin and biology, and both are required for a functioning lymphatic system. 8
Distal wing bud grafting experiments of chick into quail embryos have been shown to generate lymphatics comprised of both chick and quail LECs. This finding suggests that wing bud lymphatics do not exclusively develop from sprouts from nearby lymph sacs but also develop by recruiting circulating “lymphangioblasts.” 7 Macrophages or circulating endothelial progenitor cells have also been proposed as a potential nonendothelial LEC progenitor. 8 , 10 Macrophages have been observed to promote angiogenesis and act as potential sources of LECs in tumor models. 10 These tumor-associated macrophages have been shown to acquire LEC-specific markers, such as prospero homeobox 1 (PROX1), vascular endothelial growth factor receptor 3 (VEGFR-3), lymphatic vessel endothelial hyaluronan receptor 1 (LYVE1), and podoplanin while simultaneously undergoing observable downregulation of myeloid markers. 7 Such “reprogramming” of myeloid into LEC identity suggests that tumor-associated macrophages and possibly other macrophages may contribute to the lymphatic vasculature, 8 although studies in developing mouse embryos have only shown that macrophages contribute to the modulation of lymphatic vessel caliber by regulating LEC proliferation without transdifferentiation or incorporation into vessels. 8 A key feature of embryonic lymphangiogenesis is venous transdifferentiation into LECs. 11
The maintenance of LEC identity requires ongoing PROX1 expression, without which reversal to a blood endothelial phenotype occurs. 12 , 13 In contrast, negative regulatory control of lymphangiogenesis is mediated by highly preserved mechanisms involving Notch, CYP26B1 (a retinoic acid degradation enzyme), and transforming growth factor-beta (TGF-beta)/bone morphogenic protein (BMP) signaling pathways. 14 , 15 During development, Notch expression (particularly NOTCH1) is integral to restricting the transition to LEC fate to certain areas of precursor veins. 16 , 17 During adulthood, another protein growth factor commonly implicated in fibrosis, TGF-beta inhibits LEC proliferation, 10 , 18 as have BMP2, BMP4, and BMP. 10 TGF-beta is highly expressed in scar tissue and therefore may have a role in the prevention of lymphatic regeneration after surgery. 5 , 18
Once established, lymphatic vessels may undergo significant remodeling controlled by expression of key molecular markers with specificity for LECs, such as VEGFR-3, LYVE1, podoplanin, and NRP2, which allow LECs to acquire responsiveness to protein growth factors that are vital to forming and maturing a vascular plexus. 3 , 7 , 10 The critical signaling pathways determining LEC remodeling revolve around the vascular endothelial growth factor C (VEGF-C)/collagen and calcium-binding EGF domain-containing protein1 (CCBE1)/VEGFR-3 pathway, a signaling axis that promotes LEC migration, proliferation, and survival and thus is indispensable for both embryonic and adult lymphangiogenesis. 7 VEGF-C loss of function experiments reveal that an absence of this key growth factor results in unaltered LEC specification, because the embryos fail to form a complete lymphatic vasculature as a result of defective lymphatic sprouting. Furthermore, VEGF-C was not active in all the areas of the embryos in which it was expressed, because its ability to activate VEGFR-3 relies on VEGF-C modulation by CCBE1, which processes VEGF-C by means of a disintegrin and metalloproteinase with a thrombospondin motif (ADAMTS3)–dependent mechanism. 7 , 10
Tumor-Induced Lymphangiogenesis: Lessons for Lymphatic Remodeling
Tumors spread either by hematogenous or lymphatic spread. 3 , 7 The formation of lymphatics in both human tumors and animal models has been linked to enhanced lymphatic metastasis. Therefore the process of lymphangiogenesis has also become a target for therapeutic restriction of tumor metastasis. 3 , 7 , 19 The intense research in this area has created new knowledge regarding lymphatic growth and remodeling. Although primarily directed at inhibiting lymphatic growth, it may also generate new insights into how lymphatic regeneration may be achieved. An important mechanism contributing to tumor-induced lymphangiogenesis is outsprouting from vessels in and around a primary tumor mass from preexisting initial lymphatics. 19 As this occurs in response to stimuli that may include VEGF-C, vascular endothelial growth factor D (VEGF-D), plateletderived growth factor subunit beta (PDGFB), fibroblast growth factor 2 (FGF2), and angiopoietin, 7 each of these signaling molecules has consequently become considered potential targets for molecular therapeutics to restrict tumor-induced lymphangiogenesis. 3 The same molecules may also be targets for lymphatic regeneration. 20
Small-caliber initial lymphatic vessels and larger collecting vessels that make up the lymphatic system differ in their morphology, structure, and anatomic locations and perform distinct specialized functions that contribute to their respective normal physiologic roles. 3 , 7 , 19 Most research investigating mechanisms behind lymphogenous metastasis of cancer cells has focused on elucidating the influences of lymphangiogenic growth factors on the initial lymphatic vessel subtypes within or around a primary tumor, 19 , 21 whereas the collecting lymphatic vessels that drain the tissues of the tumor toward the regional lymph nodes have remained largely ignored. 22 Whereas ambient smaller lymphatics largely respond to lymphangiogenic factors by proliferation and/or sprouting to generate tumor-associated lymphangiogenesis, the larger collecting vessels respond to the same lymphangiogenic stimuli in a unique and quite distinctive manner. 22 – 25 Far from passive conduits of metastatic cells, more recent findings suggest that collecting lymphatic vessels undergo a significant remodeling of their own, which contributes critically to the process of tumor spread 22 , 23 (Fig. 47-1). Observations of murine models of VEGF-C-overexpressing metastatic tumors demonstrated that drainage from the primary tumor by the collecting lymphatic performs an active role in enhancing tumor dissemination through increasing fluid flow by dilating, an increase in collecting lymphatic diameter attributed to LEC proliferation. 22 – 26 Similarly, it was shown that VEGF-D secreted by a flank xenograft tumor model also induced dilation of the collecting lymphatics draining from the primary tumor to the axilla, which was critical for cancer spread to the sentinel lymph node. 22 However, in contrast to the VEGF-C model and the effect of VEGF-D on the initial lymphatics, 6 the mechanism by which circumferential dilation occurs is not by endothelial proliferation but through specific prostaglandin-mediated responses to VEGF-D. 22 Treatment with nonsteroidal antiinflammatory drugs, including aspirin, not only reverses the VEGF-D-driven morphologic remodeling of collecting lymphatics but also reduces the rate of tumor metastasis to draining lymph nodes and distant organs. 22 These studies illustrate the complexity of lymphangiogenic signaling and that molecular stimulation of lymphangiogenesis is context specific and modulated by other growth factors and drug treatment. They also highlight the need to consider the growth and maintenance of collecting lymphatics and not just the initial lymphatics in lymphedema treatment. 7
Related posts:
 CHAPTER 48 Establishment of a Lymphedema Framework
CHAPTER 48 Establishment of a Lymphedema Framework
 CHAPTER 46 Animal Models of Lymphedema
CHAPTER 3 Quality of Life Measurement Instruments
CHAPTER 18 Lymphedema Risk Factors in Breast Cancer
CHAPTER 46 Animal Models of Lymphedema
CHAPTER 3 Quality of Life Measurement Instruments
CHAPTER 18 Lymphedema Risk Factors in Breast Cancer
 CHAPTER 33 Multiple Lymphaticovenous Anastomoses and Multiple Lymphatic-Venous-Lymphatic Anastomoses
CHAPTER 33 Multiple Lymphaticovenous Anastomoses and Multiple Lymphatic-Venous-Lymphatic Anastomoses
 CHAPTER 43 Lymphedema Complications and Their Treatment
CHAPTER 43 Lymphedema Complications and Their Treatment
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


