CHAPTER 22 Biomarkers
KEY POINTS
Although the ideal biomarker showing the complete lymphatic vascular system has not yet been discovered, a combination of markers can be used to reliably detect the lymphatics.
In comparison with the blood vascular tree, there is heterogeneity of lymphatic endothelial cells in the lymphatic vascular network.
The most constant marker to detect lymphatic vessels in human tissues appears to be double staining with antibodies against CD31 and PROX1.
It is highly likely that the lymphatic vascular system was already known to Hippocrates of Kos (about 460 to 377 BC). The Alexandrian school (about 300 BC to 600 AD) called the lymphatic vessels ductus lactei. 1 Gasparo Asellius 2 began the “reinvestigation” of the lymphatics in dogs, and Jean Pecquet 3 and Olof Rudbeck 4 followed soon thereafter in humans. Three hundred years later, the invention of the electron microscope clarified the cellular nature of the delicate wall of the initial lymphatics. 5 However, efficient biomedical and cellular studies on the development and function of the lymphatics were hampered by the lack of reliable biomarkers of lymphatic endothelial cells (LECs) in health and disease. The first molecular LEC markers were found more or less by chance. However, they were the prerequisite to distinguish between LECs and blood vascular endothelial cells (BECs), and they enabled the purification of LECs in vitro and their subsequent global characterization at RNA and protein levels. Recent studies show the heterogeneity of LECs in the lymphatic vascular tree (initial lymphatics, collectors, lymph nodes, and trunks) and the organ-specific behavior of lymphatics. Therefore the ideal marker showing the complete lymphatic vascular system and differentiating it from blood vessels has not yet been found. Nevertheless, a combination of markers can be used to reliably detect the lymphatics. In this chapter we describe the molecules that have been used most widely to characterize LECs.
Ecto-5′-Nucleotidase
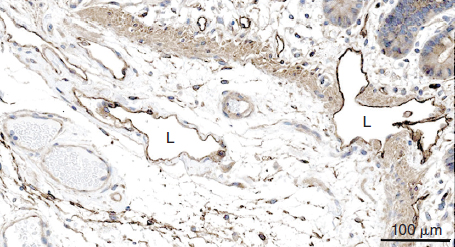
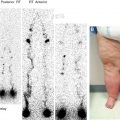
Enzyme histochemical staining for ecto-5′-nucleotidase (NT5E, cluster of differentiation 73 [CD73]) was one of the first markers to identify the lymphatics, and staining for alkaline phosphatase activity was used in parallel to identify blood vessels. 6 Ecto-5′-nucleotidase is a glycosylphosphatidylinositol-anchored surface protein, which, in addition to other functions, catalyzes the hydrolysis of purine 5′-nucleotide to nucleoside and orthophosphate in water. It is part of the cascade that completely hydrolyzes extracellular adenosine triphosphate to adenosine. It has anticoagulant activity and can be found in snake venom. 7 However, NT5E and alkaline phosphatase are not specifically expressed in LECs or BECs, and the method relies on quantitative rather than qualitative differences. Immunostaining for NT5E confirms expression in both types of vessels, with more prominent signals in the lymphatics (Fig. 22-1). Nevertheless, in combination with other techniques, NT5E can be a useful marker of the lymphatics.
CD31 and PROX1 Double Staining
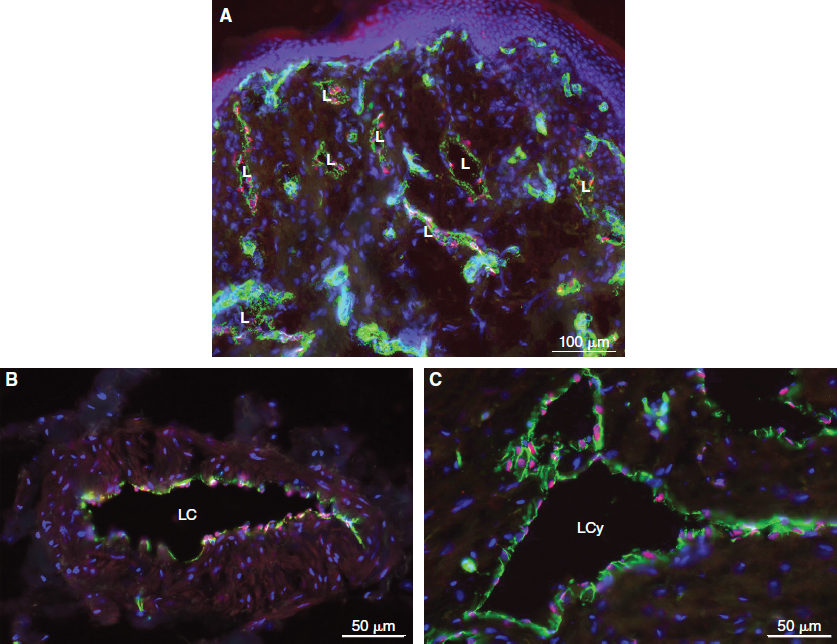
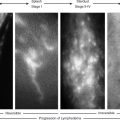
Double immunostaining with antibodies against cluster of differentiation 31 (CD31) (platelet endothelial cell adhesion molecule 1 [PECAM1]) and PROX1 is the most reliable tool to detect lymphatic vessels in human tissues (Fig. 22-2). This method detects lymphatics in human fetuses and the tissues of healthy adults, including aged adults and patients with lymphedema. It also differentiates lymphangiomas from hemangiomas, 8 although each single marker is not specifically expressed in LECs only.
PECAM1 is an adhesion molecule of the immunoglobulin superfamily. It is located in the cell membrane of platelets, monocytes, neutrophils and the subsets of T cells. 9 PECAM1 is highly enriched on the surface of BECs and to a lesser extent is expressed on LECs. It is a regulator of leukocyte diapedesis, but its functions are not completely understood, because it may have both proinflammatory and antiinflammatory functions. 10
The homeobox-containing transcription factor Prox1 is the mammalian homolog of the Drosophila gene prospero. Prox1 is essential for the development of the lymphatic vascular system. In Prox1 knockout mice, the lymphatics do not develop, whereas the blood vessels are obviously normal. 11 Because of multiple defects, the mice die during the early stages of development.
In LECs, PROX1 is located in the nucleus and thereby facilitates the counting of LEC numbers in histologic sections. Costaining with CD31 must be performed, because PROX1 is not endothelial cell specific but is expressed in other cell types, including hepatocytes, pancreatic epithelium, cardiomyocytes, lens, retina, spinal, and vegetative ganglia. 12 , 13 PROX1 is expressed in LECs but not in BECs. The only exception from this rule seems to be the valve-forming endothelial cells in the blood vascular system: the concave side of the cardiac valves and venous valves. 12 , 14 Nevertheless, the combination of CD31 and PROX1 is an excellent marker for LECs in vivo both in the initial lymphatics and lymphatic collectors (see Fig. 22-2) and in vitro.
Hyaluronan Receptor LYVE1
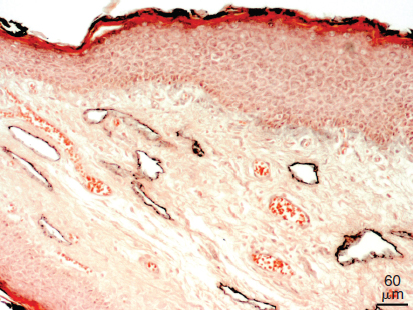
LYVE1 is a transmembrane glycoprotein and a homolog of the hyaluronan receptor CD44. 15 It is highly expressed in the initial lymphatics of the skin and other organs (Fig. 22-3) but inconsistently expressed in lymphatic collectors and lymph node sinusoids. 16 , 17 In our experience LYVE1 is an excellent marker of the lymphatics in the mouse, but in the human we have observed weak staining also in veins. Other BECs that express LYVE1 are the sinusoids of the liver and spleen, high endothelial venules, and the syncytiotrophoblast of the placenta. 18 , 19 A large number of scattered mesenchymal cells, many of which are obviously macrophages, are positive for LYVE1. 20 It has been suggested that the physiologic function of LYVE1 resides in the turnover of hyaluronan. Hyaluronan is an important component of the extracellular matrix and is abundantly expressed in the dermis. Its main degradation sites are the lymph nodes and liver, and its transport to these organs is obviously by the lymphatic vessels. 18 However, no specific alterations of the hyaluronan content were found in Lyve1 knockout mice. 21
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


