1 Anatomy and biomechanics of the hand
This chapter has attempted to preserve the character and style of the original edition by master anatomist, Robert A Chase MD.1 In addition, we have updated the text with more recent anatomic findings, clinical pearls, basic concepts in biomechanics, and full-color illustrations.
Synopsis






Introduction
During the Renaissance, Vesalius corrected early misconceptions and brought gross anatomy into proper focus. Since that time, many investigators have embellished the basic structural studies with functional, physiologic, and philosophical observations. The forearm and hand have been prominently included in those observations. Sir Charles Bell (1834),2 in his thought-provoking volume, The Hand – Its Mechanism and Vital Endowments as Evincing Design, presented a concept of hand anatomy that places it in proper context with the position of humans in the animal kingdom. Frederick Wood-Jones (1920) probed more extensively into comparative anatomy and anthropology in his excellent work, The Principles of Anatomy as Seen in the Hand.3 Duchenne (1867) carried out detailed analysis of muscular function by isolated electrical stimulation, described in his classic volume, Physiologie des Mouvements.4
Allen B Kanavel (1925) published his monograph, Infections of the Hand, which reported detailed analysis of the spaces and synovial sheaths.5 Surgery of the Hand by Sterling Bunnell (1944) became an indispensable reference during World War II.6 Emanuel B Kaplan (1953) produced the nicely illustrated, detailed volume, Functional and Surgical Anatomy of the Hand.7 Detailed studies of the integration of the intrinsic and extrinsic muscles operating the polyarticular digits may be found in the work of Landsmeer,8–11 Kaplan,12 Eyler and Markee,13 Stack,14 Tubiana and Valentin,15 and others. More recently, newer flaps intrinsic to the hand and upper extremity have been developed from more detailed investigation into vascular anatomy.16,17 Lastly, Berger,18 Viegas et al.,19 and others have expanded our knowledge of the ligamentous anatomy of the wrist.
Skin, subcutaneous tissue, and fascia
Palmar skin, in contrast, is characterized by a thick dermal layer and a heavily cornified epithelial surface. The skin is not as pliable as dorsal skin, and it is held tightly to the thick fibrous palmar fascia by diffusely distributed vertical fibers between the fascia and dermis. Stability of palmar skin is critical to hand function. At the same time, if scar fixation or loss of elasticity occurs in palmar skin, contractures and functional loss result. The skin of the palm is laden with a high concentration of specialized sensory end organs and sweat glands. The surgeon must understand the relationship of the palmar skin creases and the underlying joints in order to plan precise placement of skin incisions for exposure of joints and their related structures (Box 1.1 and Fig. 1.1).
Box 1.1 Clinical pearl
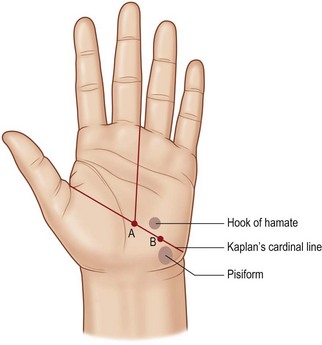
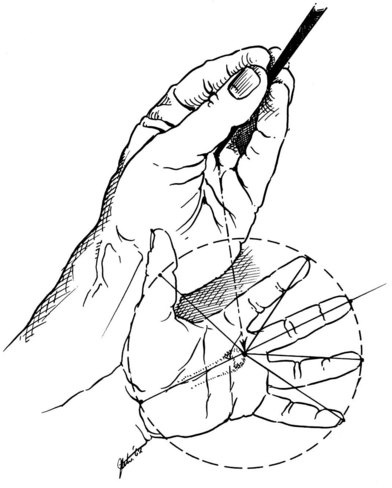
Kaplan’s cardinal line
Hand anatomist Emanuel Kaplan described specific surface lines that would aid surgeons in locating key structures in the palm of the hand. The cardinal line has often been misquoted; therefore we refer to Kaplan’s classic hand text: Functional and Surgical Anatomy of the Hand.20 Kaplan’s cardinal line is drawn from the apex of the first web space to the distal edge of the pisiform bone (Fig. 1.1). Two longitudinal lines are drawn from the ulnar aspect of the middle finger and the ulnar aspect of the ring finger. These will cross the cardinal line. The intersection of the cardinal line and the longitudinal line from the ulnar side of the middle finger corresponds to the motor branch of the median nerve. The intersection of the cardinal line and the longitudinal line from the ulnar side of the ring finger corresponds to the hook of the hamate. The motor branch of the ulnar nerve is found on the cardinal line, equidistant between the hamate and pisiform. See Kaplan’s original text for additional surface markings.
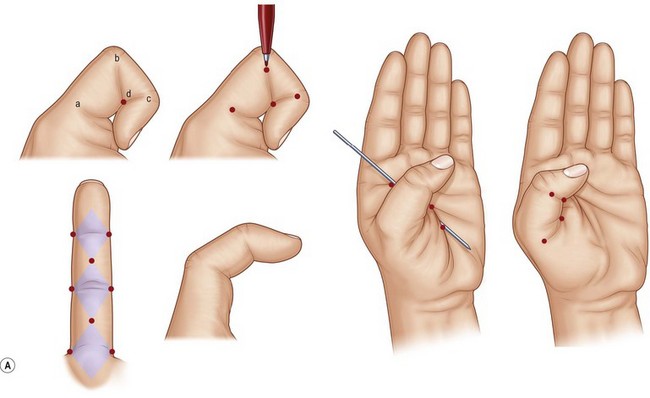
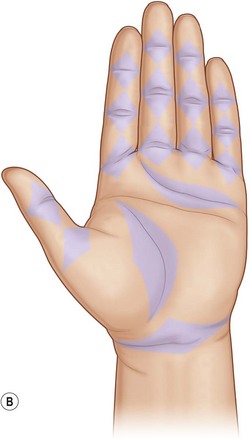
Examination of hand skin during normal ranges of motion in various planes is important in planning incisions or geometrically rearranging lacerations that might result in disabling scar contractures. Most loss of elasticity and some longitudinal shortening are compensated for adequately by mobility and elasticity of the uninjured dorsal skin. On the palmar aspect, however, scar shortening and inelasticity of the skin may result in contracture. The nature of palmar skin, its stabilizing fixation to the palmar fascia, and its position on the concave side of the hand are the bases for such contractures. Littler outlined the specific sites in the palm where a longitudinal scar would impede extension.21 For example, in each digit the geometry has been worked out by noting each joint axis and the kissing surfaces of the palmar skin in full flexion. These diamond-shaped skin surfaces should not be shortened and rendered inelastic by longitudinal scars if limitation of extension is to be avoided (Fig. 1.2).
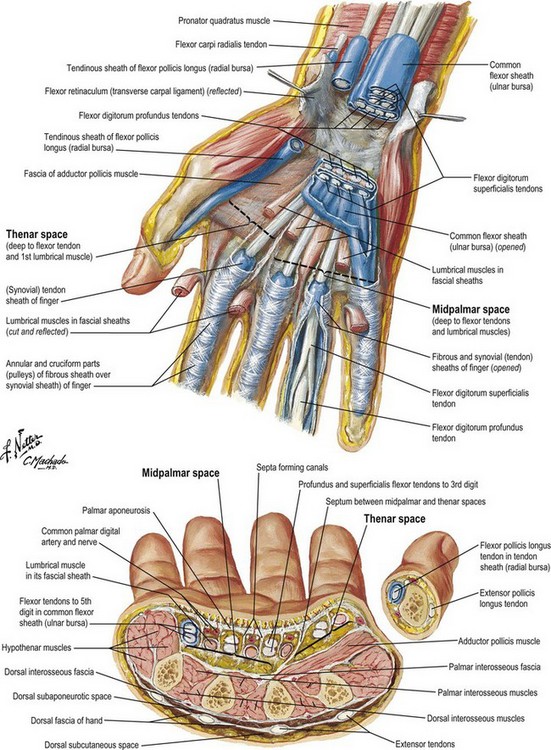
The palmar fascia consists of resistant fibrous tissue arranged in longitudinal, transverse, oblique, and vertical fibers (Fig. 1.3). The longitudinal fibers concentrate at the proximal origin of the palmar fascia at the wrist, taking origin from the palmaris longus when it is present (in about 80–85% of individuals). The fascia at this level is separable from the underlying flexor retinaculum/carpal ligament, being identified by the longitudinal orientation of its fibers in contrast to the transverse fibers of the retinaculum. The palmar fascia fibers fan out from this origin, concentrating in flat bundles to each of the digits. Generally, the fibers spread at the base of each digit and send minor fibers to the skin and the bulk of fibers distal into the fingers, where they attach to tissues making up the fibrous flexor sheath of the digits. There are attachments of the fascia to the volar plate and intermetacarpal ligaments at each side of the flexor tendon sheath at the level of the metacarpal heads.
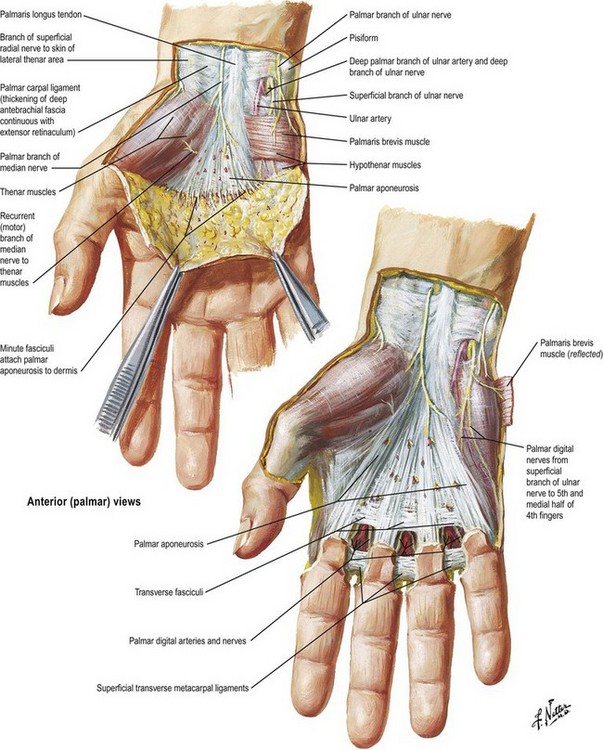
Fig. 1.3 Superficial dissection of the palm, showing orientation of the palmar fascia.
(Reprinted with permission from www.netterimages.com © Elsevier Inc. All Rights Reserved.)
Transverse fibers are concentrated in the midpalm and the web spaces. The midpalmar transverse fibers, although intimately associated with the longitudinal bundles, lie deep to them and are inseparable from the vertical fibers that concentrate into septa between the longitudinally oriented structures passing to the fingers. This system of palmar transverse fibers makes up what Skoog (1967) called the transverse palmar ligament.22 In fact, the transverse fibers form the roof of tunnels at this point that act as pulleys for the flexor tendons proximal to the level of the digital pulleys. Longitudinal fibers pass toward the palmar surface of the thumb, but these fibers are generally less numerous and sometimes difficult to identify. The thumb fibers blend into the deep fascia overlying the thenar muscles. The ulnar extreme palmar fascia blends with the hypothenar fascia. The proximal one-third of this border is the attachment site of the palmaris brevis muscle. Laterally, the muscle attaches to the hypothenar skin and hypothenar fascia.
The vertical fibers of the palmar fascia, which lie superficially to the tough triangular membrane made up by the longitudinal and transverse fibers, consist of abundant vertical fibers to the palm skin dermis (Fig. 1.4). Deep to the palmar fascia, the vertical fibers coalesce into septa, or the “perforating fibers of Legueu and Juvara,”23 forming compartments for flexor tendons to each digit and separate compartments for the neurovascular bundles together with the lumbrical muscles. There are eight such compartments, which extend proximally to about the midpalm. Proximal to this, there is a common central compartment.24 The marginal septa extend more proximally than the seven intermediate septa closing the central compartment laterally and medially. The major septum between the index flexor tendons and the neurovascular and lumbrical space to the third interspace attaches to the third metacarpal, dividing the thenar or adductor space from the midpalmar space. Knowledge of these vertical compartments aids dissection and identification of structures in operations such as trigger-finger release and Dupuytren’s fasciectomy (Fig. 1.5).
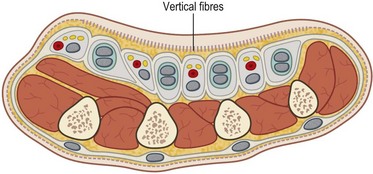
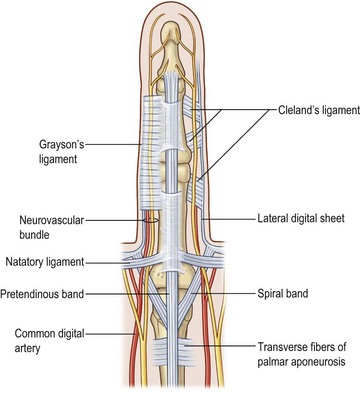
In the fingers, two important bands of fascia are named Grayson’s ligaments and Cleland’s ligaments. Grayson’s ligaments are volar to the neurovascular bundles and are quite flimsy. The much stouter Cleland’s ligaments are dorsal to the neurovascular bundles. These two fascial sheets help contain and protect the ulnar and radial digital arteries and nerves (Fig. 1.6).
Bones and joints
Hand elements
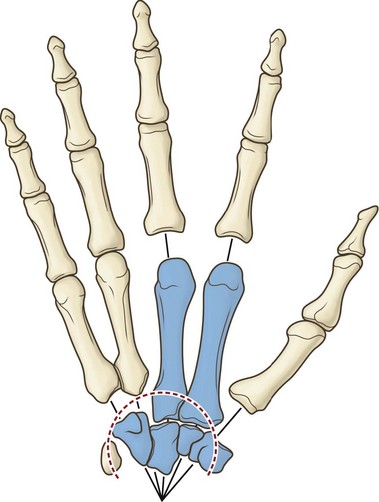
The hand skeleton is divisible into four elements:
1. The fixed unit of the hand, consisting of the second and third metacarpals and the distal carpal row.
2. The thumb and its metacarpal with a wide range of motion at the carpometacarpal joint. Five intrinsic muscles and four extrinsic muscles are specifically influential on thumb positioning and activity.
3. The index digit with independence of action within the range of motion allowed by its joints and ligaments. Three intrinsic and four extrinsic muscles allow such digital independence.
4. The third, fourth, and fifth digits with the fourth and fifth metacarpals. This unit functions as a stabilizing vise to grasp objects for manipulation by the thumb and index finger, or in concert with the other hand units in powerful grasp (Fig. 1.7).
When the head of the first metacarpal is palmar-abducted by thenar muscles innervated by the median nerve, and the fourth and fifth metacarpals are palmar-abducted by the hypothenar muscles innervated by the ulnar nerve, a volar, concave, transverse metacarpal arch is created, approximating a semicircle. The mobile metacarpal heads are pulled dorsally by extrinsic extensor tendons when the thenar and hypothenar muscles relax. It is obvious that a flaccid paralysis of the intrinsic muscles of the hand in median and ulnar nerve palsy will produce a flattened or even reversed transverse metacarpal arch. The active production of a semicircular transverse arch by the thenar and hypothenar muscles creates the proper circumferential arrangement of the metacarpophalangeal joints for convergence of the fingers in flexion. In this position the fingers, flexing at the metacarpophalangeal joints only, converge, forming with the thumb a cone, the apex of which lies over the anatomic center of the hand (Fig. 1.8). A vertical line dropped from the apex of the cone to the center of its base will strike the third metacarpophalangeal joint. This point at the apex of the transverse metacarpal arch is the anatomic center of the hand. With the fingers fully abducted, the tips form radii of equal length from the anatomic center of the hand. The same radius projected proximally falls at the wrist joint.
The wrist
The wrist joint is the site for major postural change between the arm beam and the working hand end piece (Fig. 1.9). It has a multiarticulated architecture that creates a potentially wide range of motion in flexion, extension, radial deviation, ulnar deviation, and circumduction. The distal radioulnar joint allows pronation and supination of the hand as the radius rotates around the head of the ulna. The proximal row of carpal bones (scaphoid, lunate, triquetrum, pisiform) articulates with the distal radius and ulna, providing the ability to flex and extend the hand and perform radial and ulnar deviation. The distal carpal row (trapezium, trapezoid, capitate, and hamate), along with the second and third metacarpals, forms the “fixed unit” of the hand.
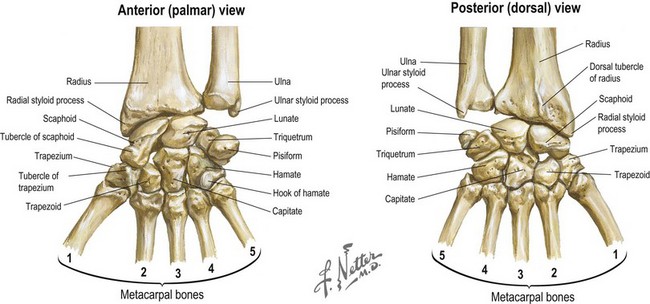
Fig. 1.9 Palmar and dorsal views of the bones of the wrist.
(Reprinted with permission from www.netterimages.com © Elsevier Inc. All Rights Reserved.)
The radiocarpal joint includes the carpal bones and the distal radius (Fig. 1.10). The principal articulation of the carpus is with the distal surface of the radius. The articular surface of the radius slopes in several planes. In the radial-to-ulnar plane, the radius exhibits an average slope of 22°. In the dorsal-to-palmar plane, the articular surface of the radius slopes 12° with the dorsal surface more distal than the palmar surface. Fractures of the distal radius frequently result in a loss of the normal radiocarpal configuration in one or both planes. A loss of the normal dorsal-to-palmar tilt of the articular surface will result in a change in the biomechanical properties of the wrist joint, which may lead to degenerative arthritis.
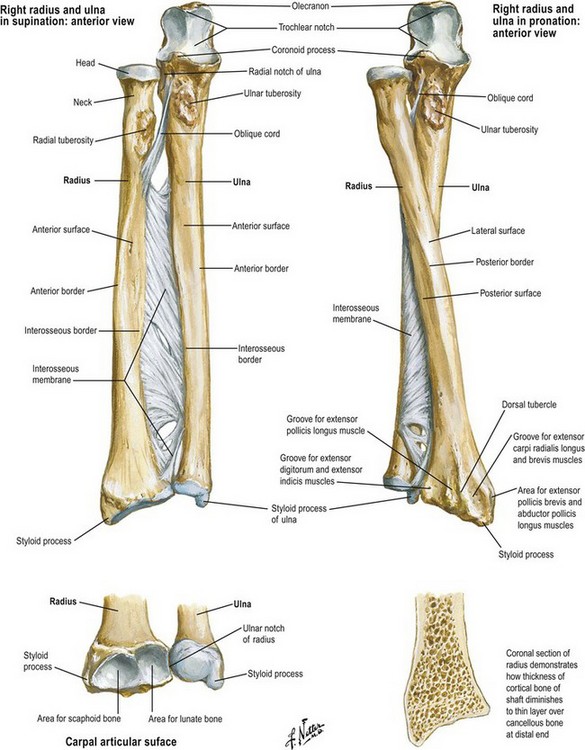
Fig. 1.10 Relationship of the radius and ulna at the proximal and distal radioulnar joints.
(Reprinted with permission from www.netterimages.com © Elsevier Inc. All Rights Reserved.)
The relationship of the length of the radius to the length of the ulna is fairly constant in individuals, and is termed ulnar variance. The distal ulna will complete the curve of the articular surface of the radius. If the end of the ulna falls short of this curvature, the condition is termed ulnar negative variance. If the ulna extends distal to this imaginary extension, the condition is termed ulnar positive variance. Either condition may lead to wrist problems. Ulnar negative variance is associated with a higher incidence of Kienbock’s disease, avascular necrosis of the lunate. Ulnar positive variance greater than 2–3 mm is associated with ulnar impaction (Fig. 1.11).
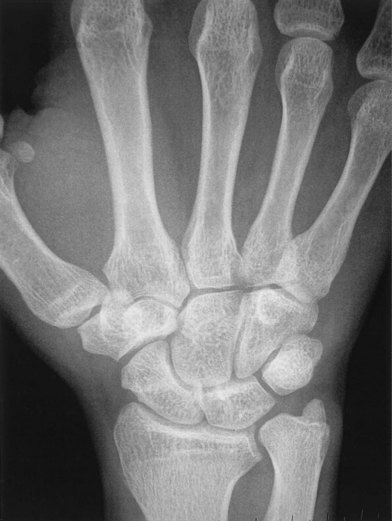
Gilula and others have described several anatomic features that denote normal extracarpal and intracarpal architecture.25 A line that follows the proximal articular contours of the proximal row of carpal bones circumscribes a smooth arc, termed the greater arc (Fig. 1.12). A disruption in the smooth appearance of this arc is one of the signs of carpal abnormality, such as abnormal rotation of one of the bones of the proximal carpal row, as would be seen with disruption of the scapholunate ligament. Similarly, the joint line between the proximal and distal row of carpal bones circumscribes another smooth arc, termed the lesser arc. The presence of abnormalities in either of these arcs may be an indication of carpal pathology, either acute or chronic.
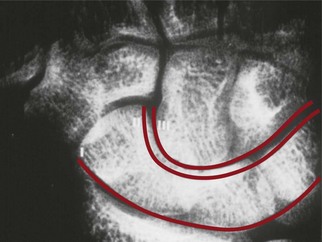
Fig. 1.12 Gilula’s lines showing the greater arc and lesser arc of the carpal bones.
(Reproduced from Hentz VR, Chase RA. Hand Surgery: A Clinical Atlas. Philadelphia: WB Saunders, 2001.)
The scaphoid and lunate bones of the proximal carpal row form the convex articular counterparts of the concave distal radius for the major wrist articulation. In fact, the articular surface of the radius is divided into scaphoid and lunate fossae (Box 1.2). The triquetrum articulates with the lunate in the proximal row, and with the hamate across the midcarpal joint. The pisiform is essentially a floating bone, unimportant for carpal stability.
Box 1.2 Clinical pearl
blood supply to the scaphoid
Investigations by Gelberman and Menon have described two main vessel systems that perfuse the scaphoid via ligamentous attachments.26 The superficial palmar branch of the radial artery contributes a volar blood supply that feeds the distal scaphoid. The dorsal carpal branch of the radial artery contributes a dorsal blood supply that also primarily feeds the distal scaphoid. Therefore, the proximal scaphoid is poorly vascularized and is susceptible to nonunion after proximal pole fracture.
All four of the bones in the distal carpal row present articular surfaces for junction with the metacarpals. The distal carpal row forms a solid architectural arch with the central capitate as the keystone. The nature of the articulations of the distal carpals with one another, and of the carpal ligament (flexor retinaculum), is such that they make up a strong and fixed transverse carpal arch (Box 1.3).
Box 1.3 Clinical pearl
checking for malrotation
The tubercle of the scaphoid is found at the distal flexion crease of the wrist joint, lateral to the tendon of the flexor carpi radialis. It is an important skeletal landmark in evaluating digital malrotations. Normally, each finger points to the scaphoid tubercle when individually flexed. A finger that points away from the tubercle may do so because of destructive flattening of the carpal arch. More commonly, it may result from malrotation following a metacarpal or phalangeal fracture (Fig. 1.13).
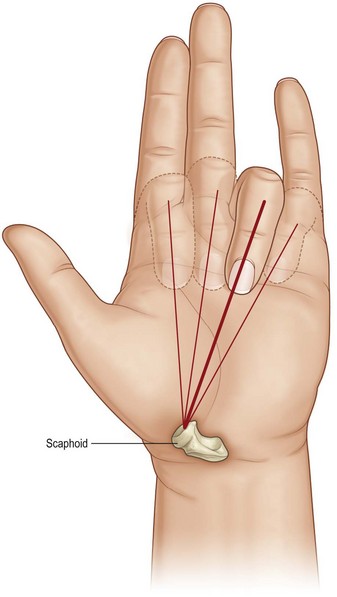
Fig. 1.13 Each finger in correct alignment points to the tubercle of the scaphoid when flexed individually.
(Redrawn after Chase RA. Atlas of Hand Surgery, vol. 1. Philadelphia: WB Saunders, 1973.)
The proximal row of carpal bones is anchored to the radius by a series of stout palmar ligaments arising primarily from the radius and by an additional set of stout ligaments arising from the ulna and the palmar portion of the triangular fibrocartilage complex. The triangular fibrocartilage complex separates the distal end of the ulna from the ulnar-sided carpal bones and serves to suspend the distal ulna to the radius at the distal radioulnar joint. These primary extrinsic palmar ligaments take the form of an inverted “V” with its apex pointed distally. Dorsally, the extrinsic radiocarpal ligament complex is thinner and is primarily a condensation of capsular tissues, except for two stout structures, the dorsal intercarpal ligament joining the distal pole of the scaphoid and the triquetrum, and the dorsal radiocarpal ligament. According to work by Viegas, these two dorsal ligaments form a unique lateral “V” configuration that allows variation in length by changing the angle of the “V” while maintaining a stabilizing force on the scaphoid during wrist range of motion.27
The intrinsic ligaments are broad, stout structures that link one carpal bone to another, either within the proximal or distal row, or linking one carpal row to the other. The two most significant intrinsic ligaments are the scapholunate ligament and the lunotriquetral ligament. The scapholunate ligament anchors the scaphoid to the lunate to allow these two carpal bones to move in synchrony. Berger has subdivided this U-shaped structure into three regions: dorsal, proximal, and palmar.28 The dorsal region is thick and controls scapholunate stability. The proximal portion, composed mainly of fibrocartilage, and the palmar region, with thin and obliquely oriented fibers, are less important for stability.29 The lunotriquetral ligament is also composed of dorsal, proximal, and palmar portions. There is less motion between these two carpal bones. Disruption of either the scapholunate or lunotriquetral ligaments may lead to wrist instability as the normal restraints on synchronous motion are removed.
Joint motion
The bony anatomy of the hand is presented in Figure 1.14. Normal metacarpophalangeal joint motion in the fingers ranges from 0 to 90°. Lateral activity in the metacarpophalangeal joints is limited by the rein-like collateral ligaments. These ligaments are loose and redundant when the metacarpophalangeal joints are in extension, allowing maximal medial and lateral deviation. As the metacarpophalangeal joint is flexed, the cam effect of the eccentrically placed ligaments and the epicondylar bowing of the collateral ligaments result in tightening and strict limitation of lateral mobility (Fig. 1.15). The fingers that have been fixed in extension during a period of healing have had the stage set for collateral ligament shrinkage and locking of the metacarpophalangeal joints in hyperextension.
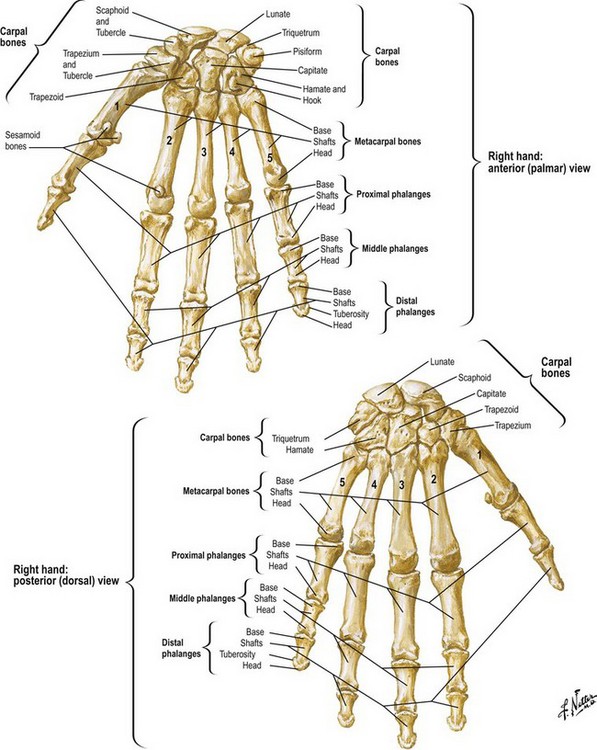
Fig. 1.14 Bony anatomy of the wrist and hand.
(Reprinted with permission from www.netterimages.com © Elsevier Inc. All Rights Reserved.)
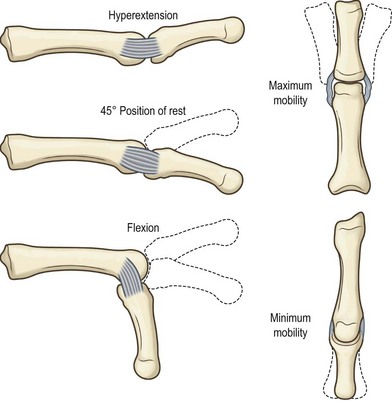
Biomechanical concept
joint motion
Brand and Hollister, in their textbook, Clinical Mechanics of the Hand, discuss how joints move.30 An axis of rotation of a joint refers to a line fixed to the proximal bone about which the motion of the distal bone appears to be a pure rotation. For the simple (hinge type) interphalangeal joints of the fingers, the motion occurs only in flexion and extension; the axis of rotation is perpendicular to the sagittal plane and is located in the distal head of the phalanx proximal to the joint. A related concept is that of the degrees of freedom of a joint. The degrees of freedom of a joint are the minimum number of axes of rotation that can be used to describe completely the motion of the bone distal to the joint. The wrist as a whole, for example, has two degrees of freedom (flexion–extension and radial–ulnar deviation), represented by two nearly perpendicular axes of rotation.31 The kinematics of more complex joints such as the thumb carpometacarpal joint32 or the intercarpal joints33 is still the subject of research and thought to have at least two degrees of freedom with nonintersecting, nonperpendicular axes of rotation.
of hyperextension. There is no lateral mobility in these joints with the collateral ligaments intact. The collateral ligaments of the distal interphalangeal joints are simply thickened medial and lateral portions of the joint capsule.
The volar plates of the metacarpophalangeal joints are the sites of insertion of the intermetacarpal ligaments, which limit separation or fanning of the metacarpal heads. The volar plate is fixed to that portion of the capsule that originates from the proximal phalanx, and therefore the plate moves with the proximal phalanx in flexion and extension. The volar plates to the proximal and distal interphalangeal joints are also stout structures that may become scarred in cases of fracture/dislocation and may ultimately lead to joint flexion contractures (Box 1.4).
Box 1.4 Clinical pearl
check rein ligaments
Check rein ligaments are, in fact, abnormal structures that are found only after fibrosis of the proximal interphalangeal joint occurs.34 The volar lateral ridges of the proximal phalanx are termed “assembly lines” and represent a coalition of the flexor sheath and Cleland’s and Grayson’s ligaments. Normally, there is no connection between the volar plate and these assembly lines. However, in the proximal interphalangeal joint with a flexion contracture, fibrosis has occurred and stout check rein “ligaments” have formed from the volar plate that prevent full joint extension.
The thumb
The thumb occupies the extreme radial position in the transverse arch of the hand.
The column of bones making up the thumb architectural base comprises the two phalanges, the metacarpal, and the trapezium. Its formula differs from that of the remaining digits by virtue of its two named phalanges rather than three. From a functional point of view, however, the thumb metacarpal can be compared to a proximal phalanx and the trapezium to a grossly foreshortened metacarpal. This suggests that the thumb is a digit recessed by a short metacarpal (the trapezium) with the proximal phalanx loosely syndactylized to the second metacarpal. There is no clear evidence to support this point of view phylogenetically, but it seems sensible in understanding the gross anatomy of the thumb’s architecture. Like the finger metacarpophalangeal joints, the thumb’s metacarpotrapezial joint has the greatest degree of freedom of any in the digital rays. The metacarpotrapezial joint is a synovial joint separated and distinct from the general intercarpal joints of the wrist. The trapezium itself articulates with the trapezoid and scaphoid with ligamentous restraints that sharply limit trapezial motion in relationship to the carpus (Fig. 1.16).
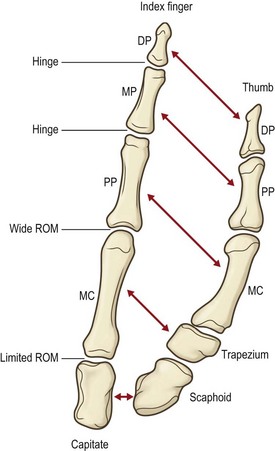
The uniquely wide range of thumb motion as compared with that of the remaining digits is attributable largely to the joint between the base of the first metacarpal and the trapezium. In simplest terms, it is described as a biconcave double saddle joint with a permissively loose capsule. That combination allows movements best described as flexion, extension, adduction, and abduction through infinite combinations that result in circumduction. The thumb rotates as a cone with its apex at a point where the axes of flexion–extension and abduction-adduction cross within the carpometacarpal joint.35–37
The first carpometacarpal joint is best described as a double saddle where one saddle sits atop another, allowing three degrees of motion: (1) flexion-extension; (2) abduction-adduction; and (3) medial rotation-lateral rotation.38

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


