7 Pathogenesis and Classification of Male Pattern Hair Loss (Androgenic Alopecia)
Summary
Keywords: androgenetic alopecia male pattern hair loss hair follicle hair miniaturization androgens testosterone 5α-dihydroxytestosterone 5α-reductase androgen receptor
Key Points
•50% of men will have developed some degree of androgenetic alopecia (AGA) by the age of 49 and around 80% of men will experience male pattern baldness by the age of 80.
•Androgens and genetic predisposition are the key factors in the pathogenesis of AGA.
•AGA can be associated with other diseases.
•The Hamilton–Norwood classification is most commonly used to describe patterns of hair loss.
7.1 Introduction
Patterned hair loss or AGA is the most common type of hair loss in men. More than 95% of hair loss problems in men are AGA. The condition is characterized by a progressive anagen shortening and premature entry into the catagen stage and a gradual replacement of strong, thick, pigmented terminal hair by fine vellus-like hair on the scalp. This hair miniaturization process occurs in certain patterns and preferentially affects hair in the temples, frontal hairline, parietal area, and vertex.
In unaffected scalp, hair has a growing anagen phase of 2 to 6 years, an involuting catagen phase of 2 to 3 weeks, and a resting telogen phase of 2 to 3 months during which some hairs are shed.1 The findings seen in AGA are a consequence of the progressively shortening anagen phases that occur in each hair cycle. Since there is less time to grow, hairs become shorter and finer. In addition, the interval between hair shedding and growth lengthens, resulting in reduced hair density.
Androgens play a central role in the process of miniaturization, hence the reason why male pattern hair loss (MPHL) is referred to as androgenic alopecia or AGA. AGA appears to result from a combination of an androgen-dependent genetic predisposition to androgen sensitivity and an androgen-independent genetic predisposition to androgen sensitivity (Video 7.1).2
7.2 Pathogenesis
Approximately 50% of men will have developed some degree of AGA by the age of 49 and around 80% of men will experience male pattern baldness by the age of 80. The incidence of pattern baldness varies from population to population and is based on genetic background. AGA shows a higher prevalence in Caucasian men compared to men of Asian or African descent.
7.2.1 Hormones and Cytokines
The influence of sex steroid hormones on the development of AGA is indisputable. The role of androgens in MPHL has been considered since ancient times. Hippocrates and Aristotle both observed the absence of baldness in eunuchs, although unsurprisingly they were unable to explain the relevant physiology. Testosterone was first implicated by James Hamilton, who observed intact hair lines in males castrated before puberty, and the cessation of hairline recession in those castrated after puberty.3 Administration of testosterone to castrati induced hair loss, which promptly stopped once testosterone was discontinued.
Testosterone has never, however, been correlated to the degree of MPHL that an individual experiences. Rather, a metabolite of testosterone, 5α-DHT , was identified as the principal causative agent.4 The metabolism of testosterone to DHT is catalyzed by the enzyme 5α-reductase, which exists as two isoenzymes: type 1 that is predominately found in sebaceous glands and the pilosebaceous apparatus; and type 2 that is present in the prostate as well as in the outer root sheath and the dermal papilla (DP) of the hair follicle. Compared with testosterone, DHT has a fivefold greater affinity for androgen receptors (ARs). The important role of DHT in MPHL is supported by both clinical and experimental evidence. Men with a genetic deficiency of type 2 5α-reductase do not develop MPHL despite normal testosterone levels. Meanwhile, studies have shown that balding scalp contains higher levels of 5α-reductase and DHT when compared to nonbalding scalp. Finally, the pharmacologic inhibitors of 5α-reductase, finasteride and dutasteride, are proven to be the most efficacious medical treatments for MPHL.
The exact molecular pathways by which DHT might cause MPHL have not been clarified. We know that androgens generate cellular responses by binding to intracellular AR and that this complex then binds to promoter DNA sequences of androgen-regulated genes, thereby altering gene expression and protein synthesis. We know that the role of AR is as important as the presence of the androgen itself, since genetically male individuals with hypofunctional AR do not develop secondary male characteristics, nor do they develop MPHL in spite of normal serum androgen levels.5 The gap in our knowledge lies in our understanding of the nature of the proteins that are expressed, and how they might enhance or suppress hair growth. Autocrine and paracrine factors are produced by balding DP cells following dihydrotestosterone (DHT)-driven alterations and may be the key factors in the pathogenesis of AGA. Hair growth may be stunted by decreases or increases of certain cytokines, such as insulin growth factor 1 (IGF-1), basic fibroblast growth factor (bFGF), vascular endothelial growth factor (VEGF), and interleukin 6 (IL-6). Insulin-like growth factor-1 (IGF-1) is known to be regulated by androgens. It was shown that DP cells in balding scalp secrete significantly less IGF-1 compared to nonbalding scalp.6 Alternatively, genetic variations may cause premature apoptosis in DP cells via the production and release of the cytokines transforming growth factor β1 (TGFβ1), interleukin 1α (IL-1α), and tumor necrosis factor α (TNFα). IL-6 is upregulated in balding DP cells compared to nonbalding DP cells. IL-6 has been shown to be secreted from balding DP cells in response to DHT.7
7.2.2 Genetic Factors
The mode of inheritance is less clear. To date, molecular genetic studies have implicated 12 genomic regions in AGA and identified a number of candidate genes.2,8,,,9,,,10,11,,,12
The most strongly associated genomic region is located on the long arm of the X- chromosome. This region is flanked by two genes, which encode for the AR and the ectodysplasin A2 receptor (EDA2R).11 This explains the maternal component of MPHL inheritance, since the gene is located on the X-chromosome. Higher expression of AR has been found in balding scalp compared to nonbalding scalp. Moreover, variations in the AR gene are associated with altered risks of developing MPHL. Men suffering from Kennedy’s disease, for example, have an abnormality of the AR gene and are highly unlikely to develop MPHL.5 Single-nucleotide polymorphism (SNP) analysis has identified still more candidate loci in the AR gene.2 The ectodysplasin A2 receptor seems to play a role in anagen maintenance. Since the X-chromosome in men is inherited from the maternal side, the association with the X-chromosome does not explain the resemblance in AGA patterns observed between affected fathers and their sons.
The remaining 11 associated regions are located on the autosomes. The strongest finding on the autosomes is for the intergenic region between paired box 1 (PAX1) and forkhead box A2 (FOXA2) on chromosome 20. Further associated regions are located on chromosomes.1,2,3,5,7,12
7.2.3 Association with Other Diseases
It was shown that early-onset vertex balding appears to be a marker for early-onset severe coronary heart disease especially in young men with hypertension or higher cholesterol levels. Matilainen et al in 2000 showed that men aged 19 to 50 who developed AGA earlier in life (<35 years of age) showed an increased incidence of hyperinsulinemia and disorders associated with insulin-resistance, such as obesity, hypertension, and dyslipidemia.13,14,15 Keum et al found an association of certain subtypes of male pattern baldness at age 45 years and colorectal neoplasia. Moreover, it was found that the incidence of prostate cancer was greater in men with male pattern baldness compared with men without AGA. On the other hand, DHT can increase the transcription of prostaglandin D2 synthase through estrogen receptor beta. An increase of PGD (2) can decrease the risk of prostate cancer and proliferation of prostate cancer cells.
The pathophysiology of the correlation between AGA and coronary heart disease, insulin-resistance, and prostate and colorectal cancer is unknown and no shared genetic factors for AGA and these associated diseases have yet been identified.
Further studies will be necessary to investigate the role of shared androgen pathways in these diseases.
7.3 Classification
Various classification systems have been developed over the past decades. Beek in 1950 evaluated 1,000 Caucasian males with patterned hair loss and classified them into two types—frontal baldness and frontovertical baldness, based on the stage of evolution. In 1951, Hamilton produced the first grading scale after examining 312 Caucasian men and 214 Caucasian women. The Hamilton scale ranges from type I to VIII. Type I represents the prepubertal scalp with terminal hair growth on the forehead and all over the scalp; type II and III show gradual frontal, mostly M-shaped recession of the hair line; type IV, V, and VI show additional gradual thinning in the vertex area; type VII and VIII show a confluence of the balding areas and leave hair only around the back and the sides of the head.3 Setty studied 300 white and 300 black males and simplified Hamilton’s classification into three different subtypes keeping into account the patterns of hair loss in black males as well. Ogata distinguished 15 different subtypes of patterned hair loss and classified them into 6 different subtypes based on the study of Japanese men. In 1975, Norwood modified the Hamilton classification, based on a study of the degree of alopecia in 1,000 Caucasian males of different ages. He included variations on the middle grades IIIa, IVa, and Va, which show a more prominent gradual receding of the middle portion of the frontal hair line and type III vertex which is characterized by a loss of hair mainly in the tonsure area and a frontotemporal recession which never exceeds that of type III. Fig. 7.1 shows the Hamilton–Norwood classification. Currently, the Hamilton–Norwood classification system is most commonly used to describe patterns of hair loss. The Hamilton–Norwood scale is used for the assessment of efficacy of drugs for hair restoration in clinical trials. However, a mixed pattern of Hamilton–Norwood types and Ludwig types often occurs in men in clinical practice.
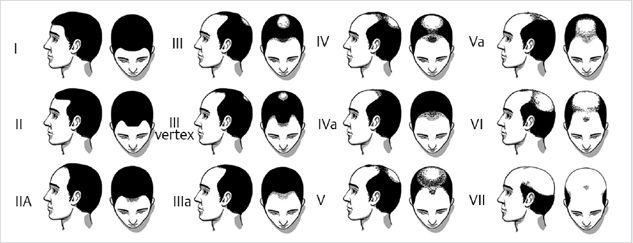
Fig. 7.1 Hamilton–Norwood classification.
Related posts:
 Over-the-Counter Preparations
Over-the-Counter Preparations
 Pathogenesis and Classification of Female Pattern Hair Loss
Pathogenesis and Classification of Female Pattern Hair Loss
 Methods and Techniques to Study Hair Cell Survival
Methods and Techniques to Study Hair Cell Survival
 Pityriasis alba
Pityriasis alba
 Cutaneous infiltrates – lymphomatous and leukemic
Cutaneous infiltrates – lymphomatous and leukemic
 Single Step Multivariant Analysis of Serum Autoantibodies in Autoimmune Blistering Diseases Using BIOCHIP® Mosaic Technology
Single Step Multivariant Analysis of Serum Autoantibodies in Autoimmune Blistering Diseases Using BIOCHIP® Mosaic Technology
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


