Key Points
- ▪
Lymphangiogenesis occurs in four distinct regulatory phases: lymphatic competence, lymphatic commitment, lymphatic speciation and lymphatic maturation.
- ▪
Vascular endothelial growth factor-C/vascular endothelial growth factor receptor-3 signaling is essential in lymphangiogenesis.
- ▪
Prospero-related homeobox-1 is a transcription factor that serves as a master regulator that specifies the differentiation of venous endothelial cells into lymphatic cells.
- ▪
Animal models of lymphedema suggest the promising role of growth factor-mediated therapies and lymph node transfer in the treatment of chronic lymphedema.
- ▪
Animal models of secondary lymphedema have expanded our understanding of its pathogenesis and have facilitated the advancement of its treatment.
- ▪
Recent reliable rodent model allows for further investigation of the physiologic mechanism, molecular basis, and long-term outcomes of vascularized lymph node transfer.
Introduction
The lymphatic system plays an essential role in maintaining tissue-fluid homeostasis and contributes to immune surveillance by trafficking of lymphocytes and antigen-presenting cells to the lymph nodes. Dysfunction of the lymphatic system results in lymphedema where regional accumulation of interstitial fluid, macromolecules, and cellular debris occurs. Long-term lymph stasis causes a chronic inflammatory response which results in skin thickening, subcutaneous tissue fibrosis and recurrent infections.
Lymphedema is a chronic and debilitating disease. In developed countries, it occurs mainly as sequelae of oncologic surgery and therapies. Currently, there is still no cure for lymphedema. Effective pharmacologic and surgical therapies remain a challenge for physicians. Research progress in this field has been limited due to lack of reliable and reproducible experimental animal models.
Overview of Lymphedema Research
The development of animal models to study the lymphatic system has resulted in research to better understand the genetic and molecular cues underlying the pathophysiology of both inherited and acquired lymphedema. These studies have demonstrated the potential use of growth factor mediated-therapies for treatment of this disease. In addition, recent successful development of animal models with chronic secondary lymphedema has made possible the refinement of surgical techniques and evidence-based assessment of treatment outcomes.
This chapter highlights the historical and current research in lymphedema providing insight into the molecular basis that characterizes the disease process. A review of the limitations and potential applications of established animal models used in research is also provided.
Molecular Investigation
Many of the advances in our understanding of the molecular mechanisms of lymphangiogenesis and disease development in the twentieth and twenty-first century have been made possible by the availability of genetic animal models. Florence Sabin described the initial theory of lymphatic development, based on experiments in pigs, as arising from primary lymph sacs that emerge from embryonic veins. Subsequent studies in genetic mouse models led to a molecular model that described early lymphatic vascular development as arising from signal-dependent differentiation of embryonic venous endothelial cells. Rockson et al. summarize the results of these earlier studies and described the progression of lymphangiogenesis as occurring in four distinct regulatory stages:
- 1.
lymphatic competence
- 2.
lymphatic commitment
- 3.
lymphatic speciation
- 4
lymphatic maturation.
Lymphatic competence is a stage between embryonic day 8.5–9.5 where all embryonic venous endothelial cells express vascular endothelial growth factor receptor-3 (VEGFR-3) and lymphatic vessel endothelial hyaluronan receptor-1 (LYVE-1) and are capable of being stimulated to undergo lymphatic differentiation. Vascular endothelial growth factor C (VEGF-C) and its receptor (VEGFR-3) have been demonstrated in many models as playing a critical role in lymphatic generation. Studies of VEGFR-3 knockout mice showed that they die early in development due to cardiovascular failure and VEGF-C knockout mice showed complete cessation of early lymphatic formation. A genetic mouse model (Chy mouse model) that expresses a heterogeneous inactivating VEGFR-3 mutation is currently used as a model for the human primary hereditary lymphedema, Milroy disease. In this Chy mouse model, lymphatic function was restored after treatment with virus-mediated delivery of recombinant VEGF-C. Crossing of the Chy mice and a mouse strain that overexpresses the VEGFR-3-specific ligand VEGF-C156S also results in restoration of lymphatic function in the double-transgenic offspring. These observations suggest the role of VEGF-C as a promising molecular therapy for treatment of lymphatic insufficiency. LYVE-1 is a lymphatic-specific cellular marker, but its role in lymphatic development is still under investigation.
Lymphatic commitment occurs at embryonic day 9.5–10.5 and is a stage where a subpopulation of embryonic venous endothelial cells is marked for commitment to lymphatic differentiation via the expression of the transcription factor, prospero-related homeobox (PROX-1). These cells are stimulated by an unknown inductive signal from the surrounding mesenchyme to express the PROX-1 transcription factor and to polarize along the cardinal vein. The role of PROX-1 as a master control of lymphatic-specific differentiation is supported by studies that demonstrated absence of lymphatic development in PROX-1 knockout mice. In addition, differentiated vascular endothelial cells that are induced to express ectopic PROX-1 can be reprogrammed to adopt a lymphatic phenotype.
Lymphatic speciation follows at embryonic day 10.5–11.5 and is characterized by expression of additional surface lymphatic-specific markers and downregulation of vascular endothelial-specific markers.
At embryonic day 11.5–12.5, rudimentary lymphatic sacs start to form from these PROX-1-positive lymphatic endothelial cells in a process called ‘lymphatic maturation’. By embryonic day 14.5, the maturation of these lymphatic vessels is nearly complete. These lymphatic structures continue to mature and organize into the adult lymphatic network, a process that continues until the early period after birth.
Animal Models
While genetic animal models have provided insights into the embryologic development of the lymphatic system and the mechanisms of primary lymphedema, animal models of secondary lymphedema have expanded our understanding of its pathogenesis and have facilitated the advancement of its treatment. Research using canine, sheep, rabbit and rodent models has reported variable success rates. The challenges of the initial studies have been to create an animal model with reproducible and sustained lymphedema.
Canine
In 1968, Olszewski et al. described a successful experimental lymphedema model in dogs by transecting the main lymphatics of the hind limb and then excising a circular strip of skin, subcutaneous tissue, fascia and periosteum, leaving the tissue gap to heal by secondary intention. This technique was modified by completely removing the lymphatic chains in the thigh in a second surgery, which resulted in an accelerated rate of lymphedema development in one-third to two-thirds of the animals. This model was subsequently used to study lymphaticovenous anastomoses. Researchers showed the limitation of this technique by showing that these anastomoses initially stay patent but eventually thrombosed because of the lymph reflux that occurred as the pressure gradient from the lymph vessel to the venule reverses when lymphedema tended to resolve.
Das et al. and Chen et al. both reported success at generating temporary lower limb lymphedema in dogs by circumferential excision of soft tissue down to the muscle fascia and then irradiating the open wound. Chen et al. further demonstrated the efficacy of vascularized lymph node transfer in treating the lymphedema, although not completely. He also showed that lympholymphatic anastomosis in microsurgical lymph node transfer has no significant clinical benefit.
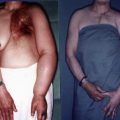
Due to the complexity of the procedures, the relatively long latency period before lymphedema is established, and the cost of maintaining a large-scale study, the canine model has not received wide acceptance in the last two decades. Recently, there has been a resurgence of interest in the canine model to study cancer-related lymphedema. Suami et al. mapped lymphatic territories in the canine forelimb and showed that there are some similarities in the deep and superficial system and size of the lymphatic vessels between canines and humans ( Figure 5.1 ). The animal model is an important milestone and allowed observations of lymphatic collateral formations after lymph node dissection in the dog, which may help to clarify understanding of lymphatic changes post lymph node dissection and radiation in humans.
Sheep
Tobbia et al. created a lymphedema model in the sheep after excision of the single lymph node in the popliteal region and ligation of the pre- and postnodal lymphatic vessels. They observed maximal postoperative lymphedema at three days after surgery, but that lymphatic function had returned by about 80% by 12–16 weeks. This demonstrates the remarkable regenerative capacity of the lymphatics and the development of collateral pathways around the site of obstruction in a large mammal. Tobbia et al. further demonstrated that vascularized lymph node transfer improves lymphatic drainage in the lower limb lymphedema that had the popliteal lymph node removed. In contrast, non-vascularized lymph node transfer did not show any improvement in lymph drainage.
Rabbit
Some researchers explored the rabbit ear as a model for lymphedema. Compared to larger mammals, this model is more affordable and surgery is less time consuming. The main neurovascular bundle and lymphatic system of the rabbit ear are readily located at the base of ear. It also lacks a deep lymphatic system. This unique anatomy of the rabbit’s ear makes surgical dissection easier. By excising a circumferential strip of tissue flap down to the chondrium and ligating the lymphatic trunk, but sparing the neurovascular bundle, Huang et al. were able to create stable lymphedema that persisted more than six months. Szuba et al. used this model to show that local administration of a human recombinant VEGF-C resulted in histologic evidence of lymphangiogenesis, improved lymphatic function and decreased dermal fibrosis compared to the untreated animals. Critics of this model were unable to replicate the same results and reported that the ear cartilage of these rabbits tended to undergo necrosis shortly after surgery. Fu et al. modified the technique by sparing the perichondrium and observed no necrosis. Using this model, they showed improved lymphatic drainage and histologic evidence of lymphatic regeneration after transplantation of lymph node fragments. A subsequent study by Yoon et al. reported that this lymphedema model was only effective in adult rabbits. Due to these difficulties in model design, researchers lost interest in this model.
In 2011, Zhou et al. described a rabbit hind limb lymphedema model that was achieved by circumferential excision of lymph nodes and deep lymphatic structure from the groin followed by removal of a skin flap from the thigh and postoperative irradiation. Applying in vitro results reported from Fang that showed that bone marrow stromal cells (BMSCs) induced VEGF-C can form lymphatic vessels, Zhou et al. showed that combined treatment of VEGF-C and BMSCs has synergistic effects on the induction of lymphangiogenesis in the treatment of acquired lymphedema in the rabbit limb model.
Rodent
The search for an inexpensive and accessible model has lead researchers to develop various models using the rodent. A model that gained initial popularity is the rat tail model. This model was popular due to the simplicity of the procedure and its effectiveness at producing lymphedema. The procedure involved performing a circumferential incision at the base of the tail and retracting the skin to create a 3 mm gap, followed by cauterization of the deep lymphatic trunk. Many investigators have used this model to clarify the role of VEGF-C/VEGFR-3 signaling and the protective role of matrix metalloproteinase (MMP)-9 in counteracting tissue swelling. Several studies have supported the observation that VEGF-C may only be useful in augmenting lymphatic proliferation in areas where lymphatics are deficient or absent, but not in cases where lymphatics are intact but poorly functional. Other studies showed that genes that are responsible for inflammation and oxidative stress are upregulated in the lymphedema rat tail model. This suggests the promising role of anti-inflammatory agents in suppressing this response.
Critics of this model claimed that it lacks correlation with human disease. The rat tail lacks lymph nodes which likely play a critical role in the development of lymphedema in humans. In addition, the hydrodynamic and cellular mechanisms of lymphatic drainage in the rat tail models are also different than in a human limb, which may influence the disease course.
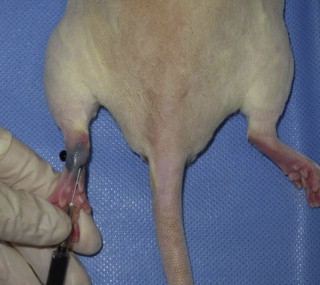
Other investigators sought to develop hind limb models. Initial models that involved removing a skin flap and resecting the underlying lymphatic trunk alone were not successful at producing sustained lower limb lymphedema due to rapid lymphatic regeneration in the rat. Wang et al. also showed an unexpected development of lymphedema in the contralateral untreated limb with this technique. Other researchers later showed that when combined with preoperative or postoperative radiation, chronic lymphedema of the hind limb can be achieved. The main disadvantage of these approaches was the high mortality rate due to the high radiation dose. Oashi et al. described a successful mouse hind limb model that combined preoperative radiation with 30 Gy, ligation of the deep lymphatics, removal of subiliac and popliteal lymph nodes, and creation of a skin gap by suturing the edges of the skin to the underlying muscle. Sommer et al. reported a technique involving just the removal the inguinal and popliteal lymph nodes and adjacent lymphatic vessels of the hind leg followed by irradiation with 15 Gy. They observed stable lymphedema at four weeks out from treatment. These same investigators also showed that regeneration of autologous transplanted lymph node fragments across the areas treated with radiation and lymphadenectomy can be achieved with exogenous injection of VEGF-C. Yang et al. further showed that chronic lymphedema of the hind limb could be achieved beyond three months by complete lymphadenectomy of the groin and popliteal region and ligation of the superficial and deep lymphatics, followed by a single dose of 20 Gy postoperatively ( Figures 5.2–5.5 ). They introduced the use of micro-computed tomography (micro-CT) reconstructed images from the ankle joint to the hip joint as a means of accurately comparing the volume differentiation between the preoperative and postoperative lymphedematous limb. They also found that skin excision was not necessary in their model. These investigators also developed a model for vascularized lymph node transfer using para-aortic lymph nodes and demonstrated that the acquired lymphedema may be corrected with vascularized lymph node transfer and that the effect is correlated with the number of lymph nodes transferred ( Figures 5.6–5.8 ).