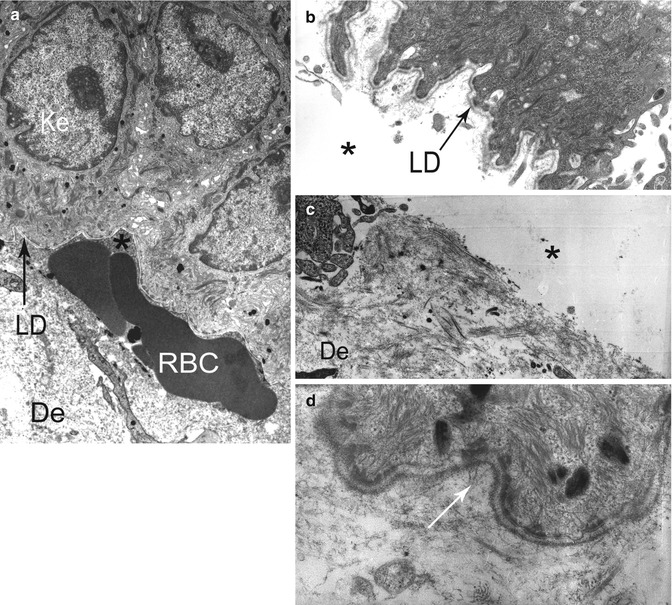
Fig. 19.1
Electron micrograph illustrating key components of a desmosome (a, b) and a normal anchoring complex (c, d): tonofilaments (tf), attachment plaque of tf in desmosomes (arrow head), hemidesmosome (HD) with inner (simple arrow) and outer plaque (double arrow), lamina lucida (LL) and lamina densa (LD), anchoring filaments (af), anchoring fibrils (AF)
Finally, standard histology and direct IF all together allow the classification of three main groups: the subepidermal AIBD, the major EBH, and the intraepidermal AIBD group of pemphigus. A fourth group of minor EBH has been recently added to the EBH classification [1]. TEM and IEM help in the differential diagnosis of types and subtypes of blistering diseases in each of these groups.
This article aims to provide the main TEM and IEM findings which are important for the differential diagnosis of blistering diseases.
19.2 Materials and Methods
19.2.1 Biopsy
Biopsies for EB and AIBD are reviewed elsewhere in this book. The minimal size of the biopsy in our opinion is 3 mm for the EM and 6 mm for the IEM. The sample must be immersed immediately in the adequate medium (see below) since any delay will cause it to dry out and result in irreversible damages, rendering it partly or totally useless for subsequent EM analysis.
19.2.2 Fixation and Specimen Processing
The basic principle of a primary and secondary fixation sequence is generally accepted as being the best method of fixing specimens for EM, while IEM asks for a specific specimen processing [2]. Typically the embedding and processing for TEM takes 4 days and for IEM takes two additional days to perform the immunolabeling. This can be achieved with horseradish peroxidase [2–6] or colloidal gold [7–10] and before [2–6, 9], after [7, 8], or without embedding [10].
19.3 Results in Individual Diseases
Information obtained by TEM and IEM are presented by disease. When the patients have multiple locations of their disease, these results are generally the same from one location to another. It should be borne in mind that these results depend on the technique used particularly in AIBD. The labeling by direct IEM reveals auto-Ab accumulated in the skin or mucous membrane of the patients and is coarser than by indirect IEM since the sera containing the auto-Ab are diluted before use. The techniques of IEM in block before embedding can only reveal membrane or extracellular antigens. Finally, the interpretation of the results has to take into consideration the ultrastructure of normal skin and mucous membranes and its variations without pathological significance. For example, a morphometric study in healthy subjects showed the number of AF is highly variable between individuals and from one region to another [11].
19.3.1 Subepidermal/Epithelial Autoimmune Bullous Diseases (AIBD)
In all subepidermal/epithelial AIBD, semithin sections show similar split and linear immune deposits at the dermoepidermal junction (DEJ) in the skin or the chorio-epithelial junction (CEJ) in the mucous membranes (Fig. 19.2). According to the subepidermal/epithelial AIBD involved, the analysis of the ultrathin sections by TEM can show a cleavage in the lamina lucida (LL) or under the lamina densa (LD) and direct IEM in vivo bound immune deposits in the LL, overlying the LD and/or below the LD. Indirect IEM allows to analyze more finely the binding sites of the circulating Ab in the anchoring complexes of DEJ or CEJ, on the HD at the basal pole of keratinocytes, on anchoring filaments in the LL beneath the HD, or on AF below the LD. Indirect IEM is contributory only in the seropositive forms of AIBD, and it does not inform about the pathogenicity of these circulating Ab. All in all, the studies using direct IEM provide diagnostic information in the seronegative forms of the subepidermal/epithelial AIBD, and the studies using indirect IEM have a physiopathological interest in the fine localization of immunodominant epitopes.
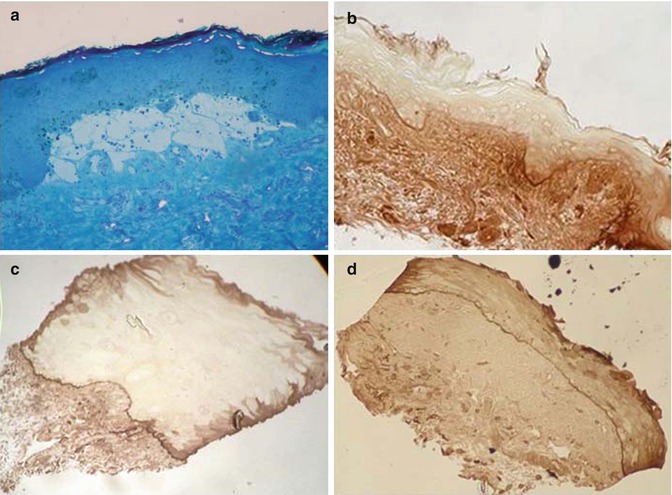
Fig. 19.2
(a) Semithin plastic-embedded sections. (a) MET technique: split between the dermis and epidermis. Toluidine blue stain (b–d) IEM technique using peroxidases: linear immune deposits at BMZ of the skin (b), buccal mucosa (c), and conjunctiva (d)—a black-brown labeling was observed, contrasting with the slight background staining
19.3.1.1 Subepidermal/Epithelial AIBD with Cleavage in the Lamina Lucida: Bullous Pemphigoid, Mucous Membrane Pemphigoid, and Pemphigoid Gestationis
The two major diseases in the group of subepidermal/epithelial AIBD with cleavage in the LL are bullous pemphigoid (BP) and mucous membrane pemphigoid (MMP), formerly called cicatricial pemphigoid (CP) [12]. In spite of their similarity, BP and CP exhibit several differences when a fine analysis by TEM is performed. The HDs at the roof of the cleavage are absent, whereas the ultrastructure of the floor is normal in BP (Fig. 19.3a–c). In contrast, the HDs along the cytoplasmic membrane of basal keratinocytes are normal, but a thickening or focal interruptions of the LD in the floor of the cleavage are observed in the CP (Fig. 19.3d) [13–15].
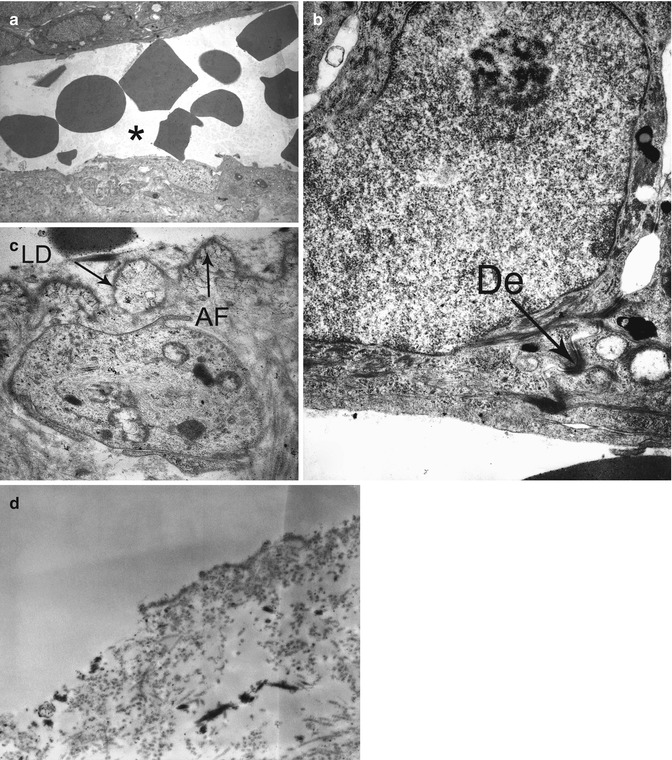
Fig. 19.3
TEM in bullous pemphigoid (a-c) and cicatricial pemphigoid (d) showing a split in the lamina lucida with red blood cells (a), absence of hemidesomomes in the roof of the cleavage in BP (b), normal lamina densa (LD) in the floor of the cleavage in BP (c), thickening and focal interruption of the LD in the floor of the cleavage in CP (d), De normal desmosomes, AF normal anchoring fibril
The studies using direct IEM allow differentiating the BP of CP in most of the cases. The first case reports with studies by direct IEM were published between 1975 and 1982 and large prospective series in 1987 and 1998 [4, 16–18]. In the skin of patients with BP, the immune deposits along unseparated zone of DEJ appear very thin. They occupy exclusively the LL and the LD is easily identified beneath them. They are very close to the plasma membrane of basal keratinocytes and stop in front of melanocytes (Fig. 19.4). Sometimes, they overflowed into the HD attachment plaque of necrotic basal keratinocytes. An infraclinical cleavage of DEJ is rarely observed in the skin of BP patients processed for IEM. If present, it is then seen in LL, and immune deposits decorate its roof (Fig. 19.5). In the skin or mucous membranes of patients with classical mucocutaneous CP, the immune deposits along unseparated zone of DEJ or CEJ appear thick, approximately two to three times the width of the deposits seen in BP. They are irregular in thickness, mostly overlying the LD and occasionally the LL (Fig. 19.6) [19]. A clear space is frequently seen between immune deposits and cytoplasmic membrane of keratinocytes. The labeling of DEJ/CEJ stops where the LD is destroyed. A subclinical split of DEJ or CEJ is frequently observed in the skin or mucous membranes of CP patients processed for IEM. It is then seen in LL, and immune deposits decorate both its roof and floor. Concordant results were obtained for testing of skin and different mucous membranes in the same patient and for ultrastructural localization of Ig and C3 in the same sample. Very few studies by direct IEM of anti-laminin-332 (LN-332) MMP (formerly called anti-epiligrin and anti-laminin 5 CP) have been published [20–22]. In this form of CP, immune deposits localized to the lower LL at its interface with the LD, exclusively to the floor of cleavage in split zone. In patients with a pure ocular CP, the localization of immune deposits in their conjunctiva may be similar to that observed in the skin of patients with a BP. Very thin deposits are observed at the upper LL region immediately subjacent to the HD (Fig. 19.7) [23]. The fineness of deposits can explain the negativity of direct IF in some patients [24]. This pattern is consistent with studies showing that the target antigen of Ab in these pure ocular CP is the chain β[beta]4 of integrin α[alpha]6β[beta]4, a transmembrane molecule of the HD [25].
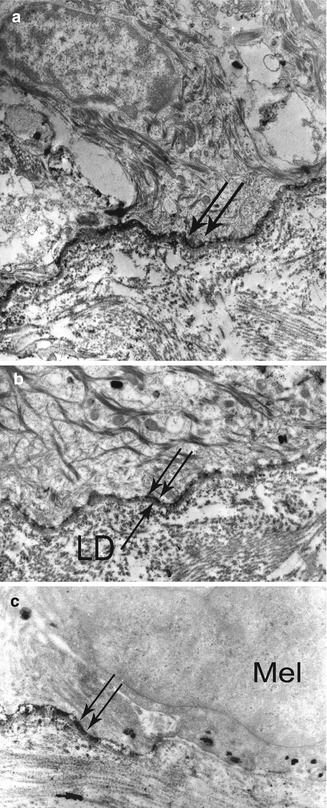
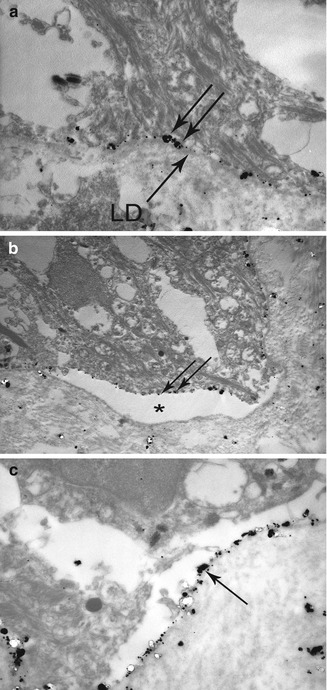
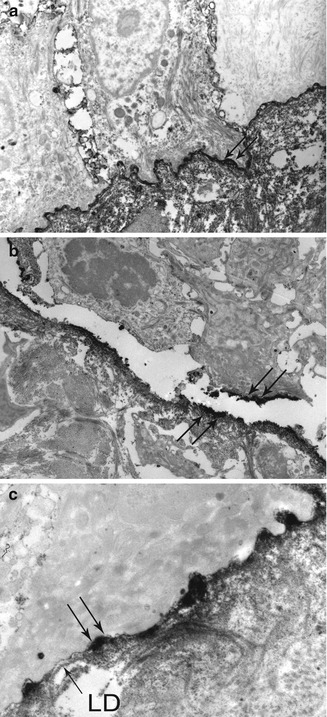
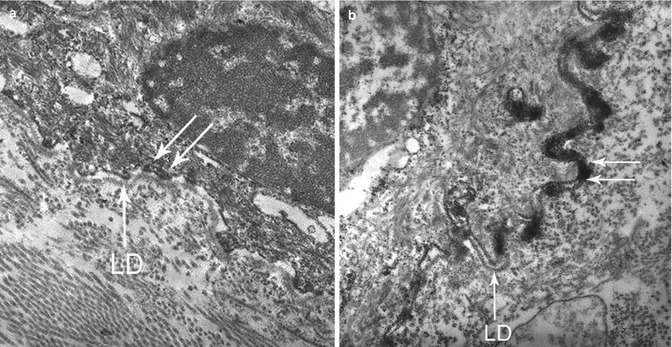
Fig. 19.4
Direct IEM using peroxidases in a BP patient: (a) very thin immune deposits (double arrow) at the dermoepidermal junction, at low magnification. (b) Immune deposits (double arrow) are exclusively located in the lamina lucida—the lamina densa (LD) is well identified below them, at high magnification. (c) Immune deposits (double arrow) stop face to melanocytes (Mel)
Fig. 19.5
Direct IEM using colloidal gold in pre-embedding technique, in a BP patient: (a) IgG deposits (double arrow) in the lamina lucida—the lamina densa (LD) is well identified below them. (b) Cleavage in the lamina lucida with IgG deposits (double arrow) at the roof. (c) Control: anti-laminin-332 monoclonal antibody GB3 deposits (->) at the floor of the cleavage
Fig. 19.6
Direct IEM in a patient with a classical mucocutaneous CP: (a) thick immune deposits (double arrow) at the dermoepidermal junction, at low magnification; (b) split in the lamina lucida with immune deposits (double arrow) both at the roof and the floor of the cleavage; (c) immune deposits (double arrow) are mostly located on the lamina densa (LD) and partly obscure the LL, in unseparated zone seen at high magnification
Fig. 19.7
Direct IEM in pure ocular cicatricial pemphigoid: immune deposits (double arrow) at the upper part of the lamina lucida (a) or overlying the lamina lucida and the lamina densa (LD) as in a mucocutaneous CP (b)
The studies using indirect IEM show also differences between the sera of patients with BP and CP (Fig. 19.8). In BP, target antigens of circulating auto-Ab are clearly associated with HD and distributed in two pools. On the one hand, there is a large intracellular pool, closely related to the attachment plaque of HD; on the other hand a small extracellular pool, in the upper lamina lucida close to HD [5, 26, 27]. More recent studies demonstrated that the first one corresponds mainly to the BP230 antigen and the second to the NC16a epitope of the BP180 antigen on which auto-Ab bind in vivo in the patient’s skin [7, 28]. In the skin of patients with BP, the intracellular BP230 antigen is not recognized by auto-Ab because large molecules such as Ig cannot penetrate the cytoplasmic membrane of keratinocytes. In the CP, the target antigens of circulating Ab are exclusively extracellular in most cases, located at the lower part of the LL and on the LD, on the anchoring filament beneath the HD, averaging 40–48 nm from the cytoplasmic membrane of keratinocytes [5, 7]. In double labeling, they colocalize with the carboxy-terminal region of BP180 antigen and LN-332. In anti-LN-332 MMP, studies by indirect IEM showed immune deposits localized to the lower LL at its interface with the LD, as in direct IEM [20]. The differences in localization patterns exhibited by BP and CP autoantibodies might well help to explain the observed clinical differences in these diseases, e.g., scarring in the CP versus non-scarring in the BP.
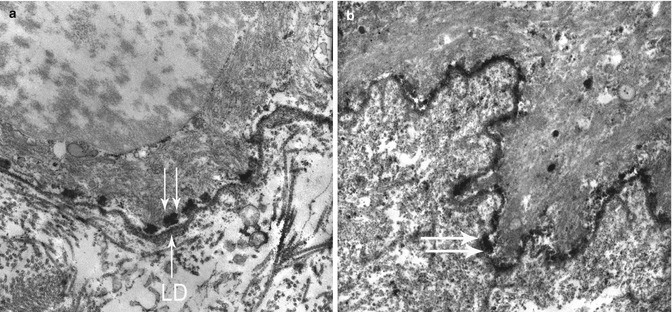
Fig. 19.8
Indirect IEM using peroxidase-labeled Ab in pre-embedding technique in BP and CP. (a) Labeling of the attachment plaque of hemidesmosomes (double arrow) by a BP patient serum, BP230+ in Western blot, after permeation with saponin—LD lamina densa. (b) Labeling of the lamina densa +/− lamina lucida (double arrow) by a CP patient serum, BP180+ in Western blot, without permeation with saponin
Pemphigoid gestationis (PG) is another, much rarer subepidermal AIBD with cleavage in the LL. It occurs during the last trimester of pregnancy and is usually regressive within 3 months after delivery. There is now evidence that it is secondary to IgG1 directed against the epitope NC16a of the BP180 antigen. Therefore, the localization of immune deposits in direct and indirect IEM is completely superimposable to that observed in the BP, namely, deposits in the upper part of the LL [29, 30].
19.3.1.2 AIBD with Cleavage Below the Lamina Densa: Epidermolysis Bullosa Acquisita and Vesiculobullous Lupus Erythematosus
The main disease in the group of subepidermal/epithelial AIBD with cleavage below the LD is epidermolysis bullosa acquisita (EBA). As early as 1979 the TEM studies in EBA showed a cleavage under the LD clearly different from that in the LL observed in the BP and CP. A careful analysis in unseparated zone also showed destruction of AF or the presence of a broad weakly electron-dense amorphous deposit (corresponding to immune deposits sufficiently intense to be seen by TEM) (Fig. 19.9) [31–33].
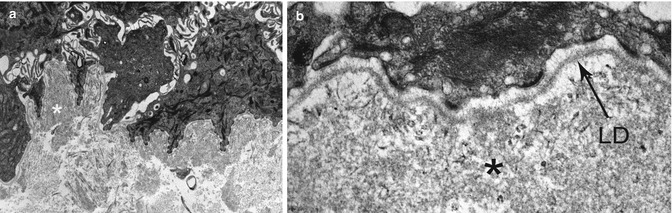
Fig. 19.9
TEM in epidermolysis bullosa acquisita showing an abnormal weakly electron-dense amorphous deposit in the AF zone, under the lamina densa (LD), at low (a) and high (b) magnification
The first case reports of EBA studied by direct IEM with peroxidase labeling were published in the 1980s [33–35] and large prospective series of 231 patients including 30 EBA and of 19 EBA in black patients of African descent later, respectively, in 1996 and 2011 [18, 36]. In the skin of patients with EBA, the immune deposits along unseparated zone of DEJ appear extremely thick. They are most often located in the AF zone, away from the LD in which they are separated by a clear space. Duplication of the LD and AF zone underlying can be observed (Fig. 19.10). Sometimes, particularly in the inflammatory form of the disease are deposits both in the AF zone and overlying the LD (Fig. 19.11). An infraclinical cleavage of DEJ is very frequently seen in the skin of EBA patients processed for IEM. It sits generally under deposits which therefore are located on the roof of cleavage (Figs. 19.10 and 19.11).
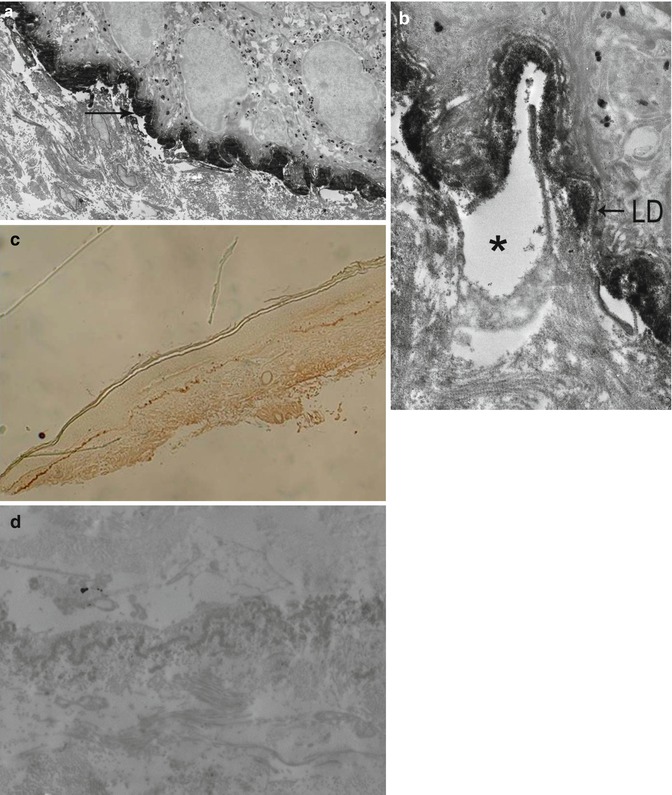
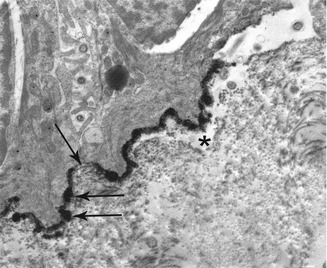
Fig. 19.10
IEM in a classical form of epidermolysis bullosa acquisita: (a) very thick immune deposits (->) at low magnification. (b) Immune deposits in the anchoring fibril zone, below the lamina densa (LD) and splits beneath them, at high magnification. (c, d) Sometimes deposits in the dermis by replication of the LD and anchoring fibril zone (c = semithin and d = ultrathin sections)
Fig. 19.11
IEM in an inflammatory form of epidermolysis bullosa acquisita showing immune deposits overlying the lamina densa (LD) and the anchoring fibril zone (double arrow) and a split beneath these immune deposits
The studies by indirect IEM with peroxidase labeling of the target antigens of circulating auto-Ab in patients with EBA give a slightly different pattern. Deposits are seen below the LD, but they only delicately decorated the AF, usually at their ends (Fig. 19.12).
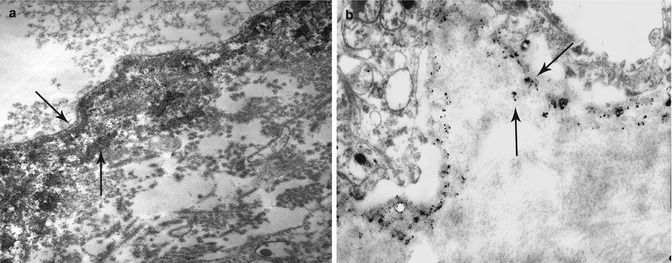
Fig. 19.12
Indirect IEM using peroxidase (a)- and colloidal gold (b)-labeled Ab in pre-embedding technique with epidermolysis bullosa acquisita patient serum, showing immune deposits that decorated the ends of anchoring fibrils
Direct and indirect IEM with gold particle studies confirm, in most patients, the labeling of AF ends that corresponds to auto-Ab directed against the type VII collagen N-terminal noncollagenous (NC) 1 domain [37–41]. However, there are discrepancies between these different studies on the precise location of AF ends. For Karpati et al. and Ishiko et al., they are localized both at the lower part of the LD and on the dermal anchoring plaques (Fig. 19.12), while for Mc Millan et al. and Ishiii et al., they are exclusively on the LD. A labeling of the central banded portion of the AF has also been reported by Tanaka in three patients whose sera reacted with the central triple-helical domain of type VII collagen [8]. Lastly, an exclusive dermal labeling has been reported by Ishii and Hashimoto [41, 42]. Ishii et al. clearly demonstrated that these patients’ sera, showing an immunoreactivity in the dermis 300–360 nm below the LD, reacted with the type VII collagen N-terminal NC2 domain.
Bullous systemic lupus erythematosus is another, much rarer subepidermal AIBD with cleavage below the LD. Patients fulfill the American College of Rheumatology diagnostic criteria for systemic lupus erythematosus and have concomitantly a vesiculobullous eruption, in relation to auto-Ab against type VII collagen [43]. Therefore, direct and indirect IEM analyses show both granular immune deposits in the superficial dermis which correspond to the large “lupus band” observed by indirect IF and linear immune deposits below the LD, as in EBA (Fig. 19.13) [44]. Sometimes immune deposits are also seen in the capillary walls.
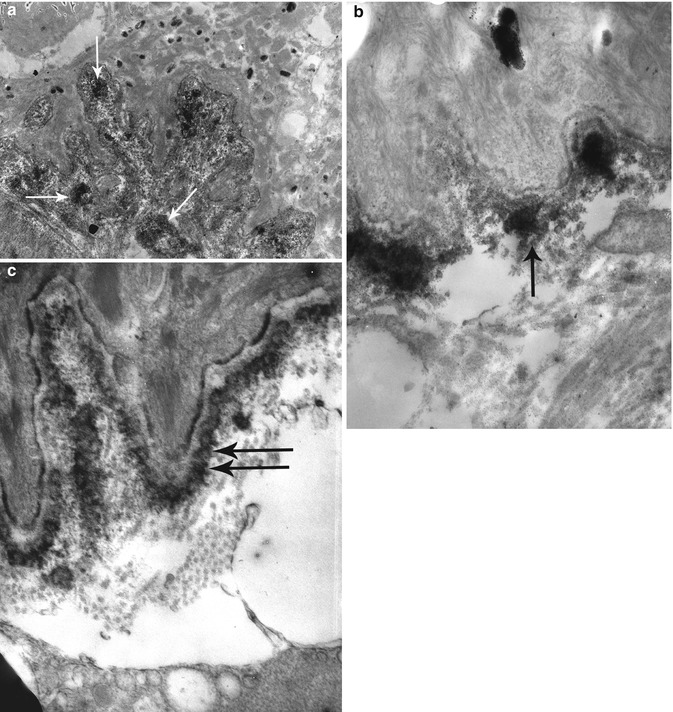
Fig. 19.13
Direct IEM in patients with systemic lupus erythematosus (LE): granular immune deposits in the superficial dermis (->) at low (a) and high (b) magnification in a patient with non-bullous LE contrasting with a linear pattern of deposits in the anchoring fibril zone (double arrow) in a patient with a bullous LE (c).
19.3.1.3 Linear IgA Bullous Diseases (LABD)
Linear IgA bullous diseases (LABD) are subepidermal/epithelial AIBD, characterized by linear deposition of IgA auto-Ab at the DEJ. They are a quite heterogeneous group under the EM. Indeed, LABD may not be a single entity but may possess target antigens shared with other subepidermal AIBD, namely, the BP, CP, and EBA [45].
Direct IEM shows many different patterns of Ab deposition in LABD. All studies have been reported before 1999 and referred by Egan et al. [45, 46]. IgA auto-Ab were localized to the LL, either on the basal surface of keratinocytes or adjacent to the HD, the LD, and the AF zone or on each side of the LD in a “mirror image” pattern, i.e., both within the LL and the sub-LD region, in respectively 26, 4, 49, and 20 % of a total of 67 patients (Fig. 19.14).
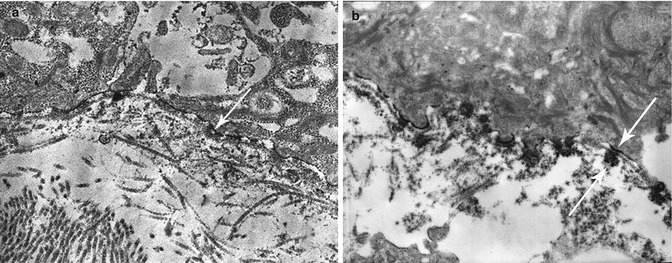
Fig. 19.14
Direct IEM in LAD: (a) immune deposits (->) in LL, (b) immune deposits (->) both in LL and below LD (mirror image)
All studies by indirect IEM in LABD have been reported between 1994 and 1998, apart from that of Bhogal in 1987 [45, 47]. They cover a total of 38 patients who were selected on the presence of a high titer of Ab in their serum. The vast majority (82 %) of these patients had sera labeling the roof of salt-split skin (SSS) by indirect IF and LL by indirect IEM [9, 10, 48, 49]. A minority (18 %) had sera labeling the floor of SSS by indirect IF and the AF zone by indirect IEM [9, 42, 47]. In one of these last cases, the serum labeled the AF dermal end but did not label the LD [42]. To our knowledge, no study by indirect IEM with LABD sera labeling both sides of SSS by indirect IF [50] has been reported.
Correlation of immunoblotting and immunogold EM studies shows that all the auto-Ab recognizing proteins in epidermal extracts of 180 kDa (or its breakdown products of 97 or 120 kDa), 200 kDa, or 230 kDa bound to the LL [9, 10, 48, 51]. The LABD97 antigen is a conformation-sensitive epitope of BP180 which needs a study by immunogold EM with cryoultramicrotomy to be demonstrated instead of cryosubstitution [10]. In contrast, auto-Ab binding to the AF zone and blotting type VII collagen has been reported in only four case reports [42, 52–54]. Indeed, dermal target antigens of auto-Ab in LABD are still only partially characterized. On the one hand, Dmochowski et al. and Egan et al. reported, respectively, 4 and 17 LABD whose sera had dermal binding of IgA on IIF and did not react with type VII collagen on immunoblot [45, 55]. Unfortunately, these sera have not been tested by indirect IEM. On the other hand, most of the patients with dermal binding of IgA on direct IEM had no circulating auto-Ab allowing indirect IEM, immunoblotting, and ELISA studies [46, 47]. These data may explain the apparent discrepancy in the LL labeling versus that of the AF zone, between direct and indirect IEM studies reported in the literature.
19.3.2 Major Types of Epidermolysis Bullosa Hereditaria
In almost all major types of EBH, the semithin sections show a cleavage along the DEJ or CEJ (Fig. 19.15). TEM can distinguish three types of major EBH which differ in the precise level of cleavage within the DEJ or CEJ: EBH simplex (EBS), junctional EBH (JEB), and dystrophic EBH (DEB) [1]. A detailed analysis of ultrastructure may show additional abnormalities of DEJ/CEJ anchoring complex allowing a diagnosis of the EBH subtype.
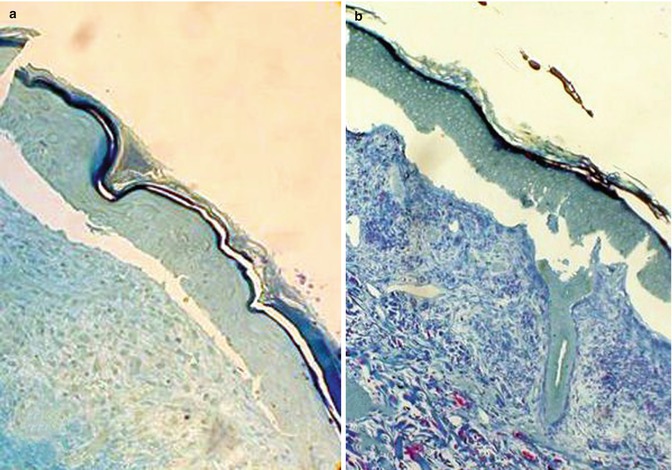
Fig. 19.15
Semithin plastic-embedded sections showing a subepidermal cleavage in EBH. Toluidine blue stain. (a) Clean split between the dermis and epidermis in a JEB. (b) Nibbled pattern of DEJ in an EBS
19.3.2.1 EB Simplex with Cleavage in Basal Keratinocytes
In most forms of EBS, the split occurs through the lower part of basal keratinocytes beneath their nuclei (Fig. 19.16). The distance, usually very small between the cleavage and BMZ itself, explains that in most cases the cleavage seems to be subepidermal/epithelial in light microscopy. Other ultrastructural changes can be observed. They are particularly useful when no cleavage is seen by TEM. In both subtypes of dominant classical EBS (localized and generalized EBS formerly called EBS Weber–Cockayne and Koebner), at first, the cytoplasm with the cell organelles appears to become diluted, and then holes develop in the cytoplasm, merge, and finally form cytolytic blisters. Although due to mutations in the genes encoding K5 or K14 keratins, the tonofilaments as well as all other constituents of the basal keratinocytes look perfectly normal [56, 57]. In contrast, in other EBS subtypes, ultrastructural abnormalities of tonofilaments can clearly provide clues to the underlying molecular defects of keratins. In generalized EBS Dowling–Meara, they can form clumps within basal keratinocytes that are due to abnormalities in the N- or C-terminal segment of the rod domain of K5 or K14 keratins, instead of the central part in classical EBS [58, 59]. In generalized autosomal recessive EBS caused by null mutations in the K14 gene, the cytoplasm in the basal keratinocytes is clarified, devoid of tonofilaments. A thin meshwork of filaments made of thin protofilaments is seen in places [60–62]. In EBS with muscular dystrophy (EBS-MD) or pyloric atresia (EBS-PA), the cleavage is located just above the HDs whose attachment plaques are small or absent. This pattern is explained by the plectin or integrin α[alpha]6β[beta]4 defects underlying this subtype of EB [63–66]. However, this pattern is not pathognomonic and may be misleading because a similar pattern has been reported in EBS without muscular dystrophy or pyloric atresia related to defect of cytoplasmic domain of integrin β[beta]4 [67] or BP180/collagen XVII [68, 69] or BPAG1-e/BP230 antigen [70].
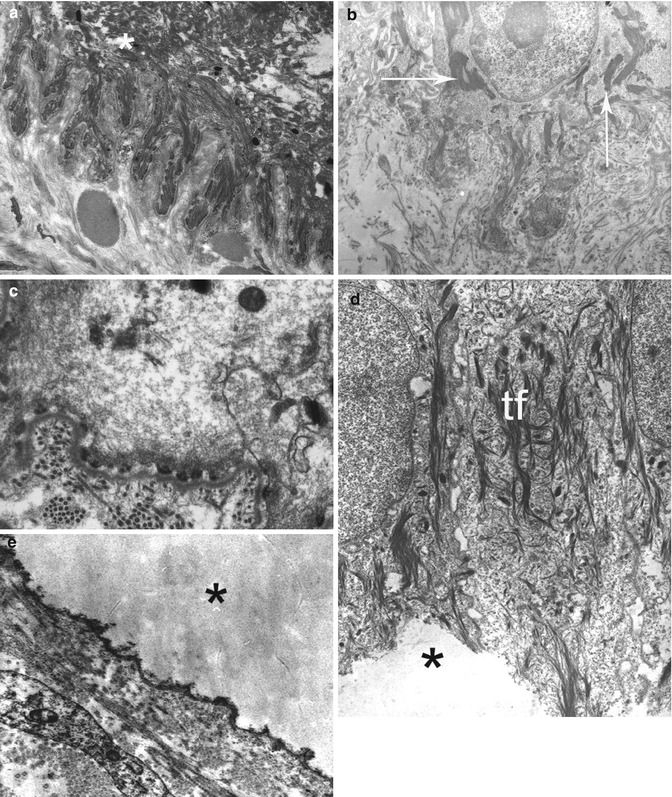
Fig. 19.16
EBS in TEM: (a, b) EBS Dowling–Meara. Note extensive split through basal keratinocyte (a) and tonofilament clumps (->) in unsplit area (b). (c) Generalized autosomal recessive EBS. A thin meshwork of filaments is seen in the cytoplasm of basal keratinocytes. (d, e) EBS with pyloric atresia. The split has occurred through the lower pole of the basal keratinocytes, leaving the cell membrane and diminutive hemidesmosome plaques with the basement membrane at the floor (->). Tonofilaments (tf) are normal
19.3.2.2 Junctional EB with Cleavage in the Lamina Lucida
All the JEB are characterized by separation through the LL of the BMZ (Fig. 19.17). Nevertheless, they display a remarkable clinical, genetic, and ultrastructural heterogeneity [71]. They include two subtypes: the very severe lethal JEB Herlitz (JEB-H) and JEB other (JEB-O) including the JEB non-Herlitz. In the first subtype, the majority of the mutations have been identified in the three genes encoding LN-332, while in the second subtype, they are usually in LN-332 or collagen XVII genes. Extracellular portion of collagen XVII and LN-332 are both components of anchoring filaments beneath HDs whose defect explains very well the cleavage in the LL. JEB-O include also some EB with pyloric atresia, named JEB-PA, that results from mutations in integrin α[alpha]6 and β[beta]4 genes [66]. A number of studies had shown qualitative and quantitative ultrastructural changes of HDs which had been initially considered as the primary defects and of diagnostic value [72–76]. In view of the current genetic data, they now appear to be secondary rather than primary. Usually HDs are sparse and small with poorly formed sub-basal plate in JEB-H, while they are normal in number and appearance in JEB-O. However, normal HDs have been reported in the JEB-H subtype and vice versa hypoplastic HDs in the JEB-O one [77–80]. Finally, an abnormal insertion of tonofilaments on the HD attachment plaque has been described in some patients [76, 81]. It may explain the extension of LL cleavage at the basal pole of keratinocytes. Therefore, even though the HD abnormalities are more frequent in the JEB-H, they do not correlate sufficiently well with the clinical outcome to be useful as a prognostic indicator.
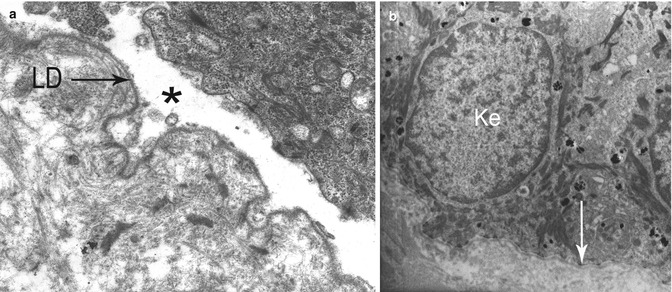
Fig. 19.17
Junctional EB (JEB) in TEM: (a) Lethal JEB Herlitz. A clean split is present through the lamina lucida, between the lower plasma membrane of a basal keratinocyte and a continuous lamina densa (LD). (b) JEB other. Unsplit skin showing small hemidesmosomes (->) at bottom surface of basal keratinocytes (Ke)
19.3.2.3 Dystrophic EB with Cleavage Below the Lamina Densa
DEB is characterized by cleavage immediately below the LD (Fig. 19.18). Several studies using EM and/or IEM and trying to correlate clinical and ultrastructural phenotypes and genotype were published before the new nomenclature of EB [1, 82–86]. All authors agree that most patients with HS-RDEB/RDEB-sev-gen have no AF in EM and no immunostaining of DEJ by IF and IEM using a monoclonal Ab (LH 7:2) that binds to the NC-1 globular domain of type VII collagen. This is explained by the absence or drastic reduction of type VII collagen synthesis, which itself is the result of mutations in the type VII collagen gene (COL7A1) leading to premature stop codons. Nevertheless, some patients with RDEB-sev-gen and normal or near-normal collagen VII immunostaining have been reported [86]. They had fewer AF which were shorter and thinner than normal, without characteristic, centrosymmetric banding pattern in EM. IEM showed no type VII collagen immunostaining or a normal labeling of both the LD and the dermal anchoring plates or a labeling restricted to the LD (Fig. 19.19). This last aspect suggests a defect in the alignment of two type VII collagen molecules into antiparallel dimers. A contrast between a quite normal type VII collagen immunostaining in IF and absent AF in EM has also been reported in RDEB-I [87, 88]. In DDEB and RDEB-O, the variability in the clinical phenotype is accompanied by heterogeneous type VII collagen immunostaining by IF and anchoring fibril morphology by EM: labeling by LH7:2 may be normal or decreased and present AF more or less rare, short, and thin (Fig. 19.18d). A detailed quantitative analysis failed to find significant differences in AF between these two subtypes [82]. In conclusion, although RDEB-sev-gen are usually recognized by the combination of negative type VII collagen immunostaining in IF and absence of AF in EM, the DDEB and RDEB-O subtypes cannot be differentiated on the basis of IF and TEM studies.
 Kindlin-1 and Its Role in Kindler Syndrome
Kindlin-1 and Its Role in Kindler Syndrome
 Cyclophosphamide in Autoimmune Blistering Diseases: Safety, Efficacy and Evidence Base
Management of Bullous Systemic Lupus Erythematosus
Cyclophosphamide in Autoimmune Blistering Diseases: Safety, Efficacy and Evidence Base
Management of Bullous Systemic Lupus Erythematosus
 Using Intravenous Immunoglobulins in Autoimmune Bullous Diseases
Using Intravenous Immunoglobulins in Autoimmune Bullous Diseases
 Living with Epidermolysis Bullosa: Reviewing the Impact on Individuals’ Quality of Life
Living with Epidermolysis Bullosa: Reviewing the Impact on Individuals’ Quality of Life
 Dermatitis Herpetiformis
Dermatitis Herpetiformis




Related posts:
Kindlin-1 and Its Role in Kindler Syndrome
Cyclophosphamide in Autoimmune Blistering Diseases: Safety, Efficacy and Evidence Base
Management of Bullous Systemic Lupus Erythematosus
Using Intravenous Immunoglobulins in Autoimmune Bullous Diseases
Living with Epidermolysis Bullosa: Reviewing the Impact on Individuals’ Quality of Life
Dermatitis Herpetiformis
Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
