Chapter 13 ADIPOSE TISSUE IN WOUND HEALING
Wound healing encompasses the varied physiologic responses of tissue following injury in an effort to restore homeostasis of the integument. This process requires precise spatial and temporal coordination of multiple intracellular and extracellular signaling pathways, the complexity of which is still being uncovered. After an injury, the blood coagulation cascade must be activated to prevent blood loss. In parallel, cytokines and other trophic mediators are released, thus recruiting circulating inflammatory and progenitor cells to the wound and stimulating tissue-resident cells. Inflammatory cells fight bacterial colonization and sequester necrotic tissue while also secreting factors that propagate the wound-healing response. Activated progenitor cells release growth factors that promote new vessel formation while modulating fibrosis and inflammation. Endothelial cells, keratinocytes, and fibroblasts react to released factors while also secreting factors to restore tissue integrity.
The complexity of wound healing is all the more impressive when considering the rarity of tissue hyperplasia and malignancy following injury. 1 Instead, injured tissue is typically replaced by a mix of fibroblasts and disorganized collagen, commonly referred to as a scar. Although scarring may serve an initially useful protective function, scars unfortunately do not retain the biomechanical properties of healthy, undamaged tissue. This is especially evident in myocardial infarction or liver cirrhosis, where the long-term effects of dysfunctional tissue healing are significant. For cutaneous wounds, scarring often fails to reproduce the mechanical and aesthetic properties of healthy skin. Burn patients, for example, often suffer disfiguring and debilitating scar contractures that limit joint mobility and function. It is therefore widely acknowledged that recapitulation of the native tissue architecture is necessary to optimize functional outcome following injury.
Interestingly, some eukaryotic organisms are capable of regenerating their original tissue architecture. Amongst mammals, early gestation fetuses are capable of regeneration, but this capacity is gradually lost during embryologic development. 2 – 4 Understanding tissue regeneration or scarless wound healing is a long-standing goal for both researchers and clinicians. To that end, a growing number of experimental therapies are being developed to promote regeneration and to minimize fibrosis, including the delivery of stem cells, growth factors, micro RNAs, small molecules or extracellular tissue scaffolds, among others. Stem cell–based therapies have already shown significant promise in preclinical trials. Adipose-derived mesenchymal stem cells (ADSCs) encompass the most promising source of stem cells for therapeutic applications, given their relative abundance and ease of harvest from lipoaspirate. 5 As a result, ADSCs have catapulted to the forefront of regenerative medicine research.
In this chapter we discuss the tissue healing capacity of ADSCs in the context of the physiologic phases of wound healing and the pathology underlying impaired wound healing, highlighting their influence on the cellular and molecular pathways governing these processes. ADSCs and wound healing encompass an active area of research. We therefore encourage readers to refer to Medline (http://www-ncbi-nlm-nih-gov.easyaccess2.lib.cuhk.edu.hk/pubmed) for the most up-to-date reviews in this field.
The Phases o f Wound Healing
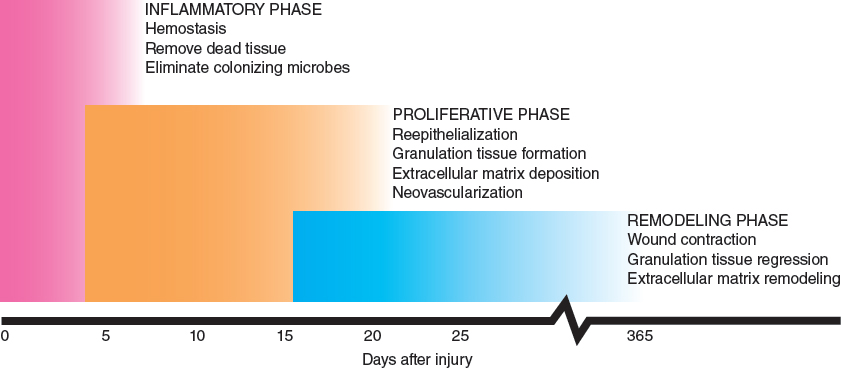
Human wound healing proceeds through three distinct but overlapping phases. During the initial inflammatory phase, hemostasis is achieved, while inflammatory cells are recruited to remove devitalized tissue and combat microbial colonization. The proliferative phase is characterized by cellular proliferation, migration, and extracellular matrix (ECM) deposition. The final remodeling phase is characterized by gradual ECM remodeling to maximize the final structural integrity of the healed wound.
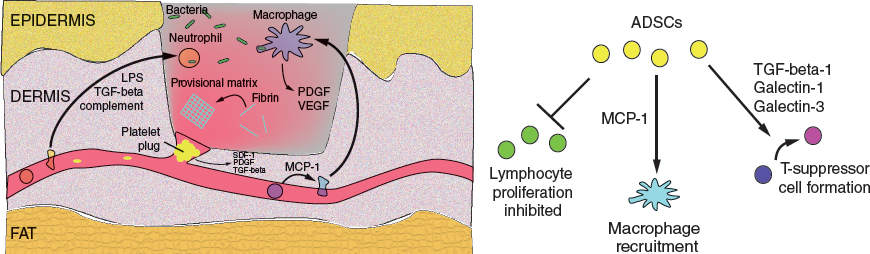
The principal processes of the inflammatory phase serve to achieve hemostasis and recruit inflammatory cells to remove devitalized tissue, eliminate microbes from the wound bed, and secrete additional soluble factors to propagate the wound-healing response. ADSCs modulate inflammation by promoting T-suppressor formation, recruiting monocytes, and inhibiting activated lymphocyte proliferation.
The inflammatory phase is initiated immediately following tissue injury. The principal goals of this phase are to achieve hemostasis, as well as to recruit inflammatory cells that remove devitalized tissue, eliminate colonizing microbes, and coordinate the ongoing wound-healing response.
During hemostasis, fibrillar collagen and tissue factor activate the coagulation cascade, ultimately resulting in the conversion of fibrinogen to fibrin, which then polymerizes to form a provisional matrix. This matrix serves as a substrate for cell recruitment and engraftment, including both inflammatory and progenitor cells. In parallel, activated platelets aggregate and plug disrupted vessels. Several signaling factors including stromal cell–derived-factor-1 (SDF-1), platelet-derived growth factor (PDGF) and transforming growth factor-beta (TGF-beta) are released by platelets to activate downstream effectors. SDF-1 in particular has a well-described role in cell recruitment and governing of stem cell behavior. 6 ADSCs are capable of secreting these and other trophic factors, supplementing a normal endogenous response or correcting an impaired response.
Inflammatory cell recruitment occurs rapidly, mediated by trophic factors including TGF-beta from degranulating platelets, complement cascade proteins, and lipopolysaccharide from degraded bacteria. 7 During the first 2 or 3 days, neutrophils are the predominant cell type present within the inflammatory infiltrate. Neutrophils phagocytose dead tissue and kill bacteria through free radical- and cell-mediated cytotoxic mechanisms. They also secrete proteases to degrade ECM and prepare the wound bed for cellular ingress. 8 While neutrophils are important for preventing acute infection, their long-term presence in wounds is associated with progression to chronic, non-healing wounds. 9 ADSCs are known to protect neighboring cells from oxidative stress, 10 and can support the viability and activity of neutrophils. 11 Thus ADSCs may help promote early microbial clearance while protecting native tissues.
Circulating monocytes, recruited to the wound largely by monocyte chemoattractant protein-1 (MCP-1), differentiate into macrophages. By 3 days after the injury, they represent the major cell type in the wound. Like neutrophils, macrophages phagocytose devitalized tissue and bacteria. In addition, macrophages produce growth factors such as PDGF and vascular endothelial growth factor (VEGF) that are important for ECM production and vasculogenesis, and studies have demonstrated their critical role in wound healing. 12 Interestingly, exogenously delivered ADSCs are known to express MCP-1, suggesting a potential mechanism for improved healing by supporting monocyte recruitment to the wound bed. 13
Five to 7 days after the injury, lymphocytes enter the wound. 14 Although the lymphocytes play a critical role in eliminating foreign microbes from the wound bed, persistent lymphocytic activation can be associated with delayed wound healing. 15 ADSC delivery may help address this problem by inhibiting the proliferation of activated lymphocytes through cell-cell interactions and paracrine signaling pathways. 16 ADSCs further promote immune tolerance by secreting TGF-beta-1 17 and galectin-1 and galectin-3, 18 paracrine factors that induce differentiation of T-helper cells into T-suppressor cells. T-suppressors positively affect wound healing, likely by inhibiting excessive inflammation while the wound matures. 19
It is clear that inflammation plays a critical role in wound healing, and ADSCs can profoundly influence this process. Recent animal studies have suggested a potential therapeutic benefit of ADSCs for treating a range of inflammatory diseases, from graft-versus-host disease (GVHD) 20 to rheumatoid arthritis 21 and colitis. 22 Although a few key soluble factors produced during the inflammatory phase have been mentioned, many other cytokines and growth factors have been shown to play important roles. The following table highlights a few of the most closely studied signaling factors implicated in the wound-healing process. Notably, all of these factors are expressed by ADSCs, underscoring their therapeutic potential for wound-healing applications.
PROLIFERATIVE PHASE
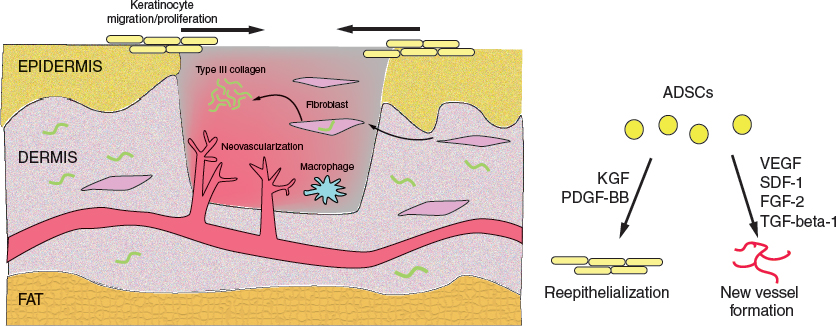
During the proliferative phase, reepithelialization occurs, while granulation tissue produces new blood vessels and type III collagen-rich ECM. ADSCs secrete multiple trophic factors that accelerate reepithelialization and promote new vessel formation.
The proliferative phase of wound healing occurs from 4 to 21 days after injury, and consists of reepithelialization, granulation tissue formation, ECM deposition and neovascularization. ADSCs are known to affect these processes through differentiation and secretion of paracrine factors. Reepithelialization likely begins immediately following injury. When keratinocytes lose their intercellular desmosomal and hemidesmosomal connections, they immediately begin migrating inward from the wound periphery via a cytoplasmic actin polymerization-dependent mechanism. Integrin receptors facilitate migration by interacting with various ECM proteins including type I collagen, fibronectin and vitronectin found within the provisional fibrin matrix. Keratinocyte migration is dependent on the production of several ECM remodeling enzymes, including matrix metalloproteinase-1 (MMP-1) and plasminogen. As cells migrate inward, peripheral keratinocytes proliferate behind the leading edge, allowing for complete wound reepithelialization. Notably, ADSCs are known to accelerate wound reepithelialization in vivo; contributory mechanisms include their secretion of keratinocyte growth factor (KGF) and PDGF-BB. 23 , 24 ADSCs have also been found to have the capacity to differentiate along epithelial lineages. 25 , 26
Four days after injury, the provisional fibrin matrix is slowly replaced by a combination of fibroblasts, macrophages, and endothelial cells, forming granulation tissue. Granulation tissue fibroblasts actively produce, deposit and remodel the ECM that comprises the healing scar while simultaneously providing a scaffold for continuing keratinocyte migration. The scar is composed mainly of the thinner type III collagen, which will eventually be replaced by thick type I collagen during the remodeling phase. TGF-beta-1 is an important regulator of ECM deposition and remodeling, and its relative paucity in fetal wounds suggests a potential contributory mechanism to scarring during adult wound healing. 27 The effect of ADSCs on fibrotic and scarring pathways will be discussed in the following section.
As the proliferative phase continues, macrophages continue to produce growth factors such as PDGF and FGF-2, which influence fibroblast behavior and promote neovascularization by inducing endothelial cell proliferation and migration. Hypoxia-inducible factor-1-alpha (HIF-1-alpha) is a master regulator of the neovascular response, and its stabilization in the setting of hypoxia results in the expression of several proangiogenic cytokines including VEGF and SDF-1. Fibroblast growth factor 2 (FGF-2), angiopoietin 1, and thrombospondin have also been shown to support angiogenesis during wound repair.
ADSCs significantly improve new blood vessel formation during the proliferative phase. The proangiogenic role of ADSCs is attributable to their expression of several key growth factors including SDF-1 and VEGF, among others. 13 , 28 – 31 Human ADSCs have been found to improve hind-limb ischemia in nude mice, 28 and in a similar model, were even shown to improve vascularity to an even greater extent than did bone marrow–derived mesenchymal stem cells (BM-MSCs), another proangiogenic stem cell population. 32 Preliminary studies of autologous ADSC therapy for critical limb ischemia in humans has shown that transplanted ADSCs are safely tolerated and lead to improved oxygenation and wound healing of peripheral tissues 33 and improvement in pain and claudication severity. 34 ADSCs are clearly able to improve blood flow and oxygenation to wounds, supporting tissue viability and accelerating wound healing.
Related posts:
 Chapter 11 THE EVOLUTION OF ADIPOSE BIOLOGY AND REGENERATIVE MEDICINE
Chapter 11 THE EVOLUTION OF ADIPOSE BIOLOGY AND REGENERATIVE MEDICINE
 Chapter 12 THE PHYSIOLOGY OF INJURED, OBESE, GRAFTED, AND DYING FAT
Chapter 12 THE PHYSIOLOGY OF INJURED, OBESE, GRAFTED, AND DYING FAT
 Chapter 14 PLATELET-RICH PLASMA AND FAT GRAFTING
Chapter 14 PLATELET-RICH PLASMA AND FAT GRAFTING
 Chapter 15 CRYOPRESERVATION OF ADIPOSE TISSUE FOR FAT GRAFTING
Chapter 15 CRYOPRESERVATION OF ADIPOSE TISSUE FOR FAT GRAFTING
 Chapter 23 SIMULTANEOUS FACELIFT AND FAT GRAFTING: COMBINED LIFTING AND FILLING FOR REJUVENATION OF THE AGING FACE
Chapter 23 SIMULTANEOUS FACELIFT AND FAT GRAFTING: COMBINED LIFTING AND FILLING FOR REJUVENATION OF THE AGING FACE
 Chapter 36 AUGMENTATION OF THE BREAST AND TUBEROUS BREAST
Chapter 36 AUGMENTATION OF THE BREAST AND TUBEROUS BREAST
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


