Fig. 1.1
Biochemical composition of dermal-epidermal basement membrane ultrastructural entities. Ultrastructural entities viewed by transmission electron microscopy (above) compared with a schematic (below) revealing their biochemical composition
Newer techniques have been developed that allow for visualization of the BMZ with greater physiological accuracy [4]. High-pressure freeze substitution allows for high-quality tissue preservation. This has illustrated that the conventional dermal-epidermal interface may in fact be inaccurate and likely an artifact secondary to tissue preparation. When tissue is dehydrated in preparation for electron microscopy fixation, the plasma membrane may be separated, exposing the lamina lucida. Therefore, the lamina lucida may be absent physiologically and the lamina densa may actually hold the anchoring filaments. From this insight, arguments about whether anchoring filament proteins reside in the lamina lucida or lamina densa become superfluous. Since conventional electron microscopy is still widely used to visualize the BMZ, conventional terms will still be used in this chapter.
The BMZ consists of complex and dynamic interactions between both intracellular and extracellular proteins, as shown in Fig. 1.2, that are important in structure, adhesion, and signal transmission. The BMZ, in addition, contains proteins that are ubiquitous throughout all tissues, as well as specialized proteins, found in particular tissues. The ubiquitous proteins, which include nidogen, type IV collagen, perlecan, and large laminins like laminin-511, form the basic scaffolding needed for basic BMZ assembly [5].
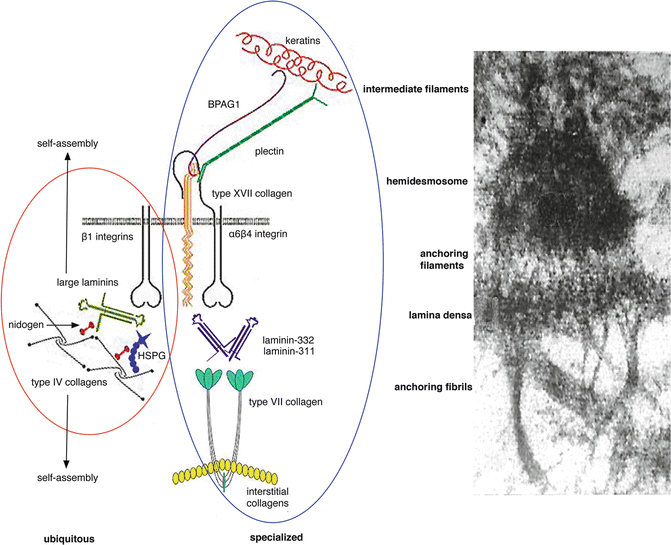
Fig. 1.2
Ubiquitous and specialized components of the dermal-epidermal basement membrane. All basement membranes (including the dermal-epidermal basement membrane) contain a group of ubiquitous components (red circle). These include large laminins such as laminin-511, type IV collagen, the heparan sulfate proteoglycan perlecan (HSPG), and nidogen. These molecules undergo a self-assembly process to form the basal lamina. In addition, stratified squamous, corneal, amniotic, and certain other epithelial tissues exposed to disruptive external forces contain a number of specialized components (blue circle) including keratins 5 and 14, keratin linker proteins BPAG1 and plectin, transmembrane proteins type XVII collagen, α[alpha]6β[beta]4 and α[alpha]3β[beta]1 integrins, specialized laminins such as laminin-332 and laminin-311, and type VII collagen. Electron microscopic image is included to the right for comparison
Newer techniques continue to illustrate the role of these ubiquitous proteins. For example, using immunogold electron microscopy and immunoblotting, perlecans were found to be key organizing proteins, connecting type IV collagen and laminins [154].
Specialized proteins are also found in tissues that are subject to external disruptive forces and need additional cohesion. Perhaps the most studied example of this type of specialized BMZ is the dermal-epidermal basement membrane zone. Starting with the most superior aspects, plectins, which are keratin linker proteins, and BPAG1, also known as BP230, are specialized intracellular proteins of the dermal-epidermal BMZ. These proteins are connected to transmembrane proteins and together form the hemidesmosome. Transmembrane proteins of the hemidesmosome include integrin α[alpha]6β[beta]4 and type XVII collagen, also known as BP180 or BPAG2. Both of these proteins also connect to extracellular proteins laminin-332 and laminin-311, together forming the anchoring filaments. The extracellular proteins also connect to the ubiquitous proteins that were discussed above to form the lamina densa. Extending perpendicular from the lamina densa are thick-banded structures, known as the anchoring fibrils. The anchoring fibrils, composed of type VII collagen, loop through the dermal interstitial collagen fibrils and reattach back onto the lamina densa. New three-dimensional human skin models are now being utilized to further investigate the BMZ [155].
The basement membrane zone is an intricate structure of ubiquitous and specialized proteins that together are key to preserving tissue structure and function. Understanding each element will illustrate the complexity and importance of the BMZ as a whole.
1.2 Ubiquitous BMZ Components
1.2.1 Nidogen/Entactin
Nidogen, also known as entactin, is a ubiquitous glycoprotein which is both a member of the BMZ and important in linking a variety of other proteins, including laminin and type IV collagen [6, 10, 11]. In fact, one of the highest binding affinities known in nature is between nidogen and laminin [10]. Two forms of nidogen have been identified: 30 mm NID1 gene-encoded nidogen-1 and 40 mm NID2 gene-encoded nidogen-2. While genetically nonhomologous, both nidogen-1 and nidogen-2 have similar structures [7]. Nidogens contain three domains, G1–G3, which are connected to each other by two rodlike segments. The G2 domain binds to type IV collagen [11], while the G3 domain binds to laminin γ[gamma]1 domain [9] and, to a lesser extent, the γ[gamma]2 chain. During development, nidogens are made by mesenchymal cells and fibroblasts in epithelial and endothelial tissue [12]. Without nidogens, epithelial tissue alone cannot form the BMZ but can with the addition of fibroblasts [13], which produce nidogen as the key factor needed to initiate BMZ production [12].
The absence of one of the two nidogens does not affect BMZ formation; however, the BMZ can be completely lost if both are absent, such as in the capillary or skin BMZ [8]. Newer studies reveal that while the absence of one form of nidogen may lead to normal histological and ultrastructural tissue with normal deposition and distribution of BMZ elements, there may still be differences in function. For example, nidogen-2- but not nidogen-1-deficient basement membranes have increased metastasis rates and larger tumors in mice lung tissue [156]. Furthermore, nidogen-2 expression has been decreased in patients with hepatocellular carcinoma [158], and nidogen-2 has been proposed as a new serum biomarker for ovarian cancer [157]. Nidogen-1-deficient mice, on the other hand, have been shown to have impaired wound healing [160]. Decreased NID1 gene expression has been demonstrated in benign nevi and primary melanoma [161]. The subtle difference in function between nidogen-1 and nidogen-2 may be due to the fact that nidogen-2 is independently binding to BMZ proteins, while nidogen-1 requires specific nidogen-binding modules found on γ[gamma]1 and γ[gamma]3 chains of laminin [159].
1.2.2 Type IV Collagen
Type IV collagen, which may comprise more than half of the mass of BMZ, is a heterotrimer containing distinct alpha chains. There are six genes that encode six different 400 nm isoforms (α[alpha]1-α[alpha]6). The isoforms have an N-terminal 7S domain (26 kDa, 28 nm), a triple helical collagenous domain (120 kDa, 320 nm) made from repeats of glycine-proline/hydroxyproline-lysine/hydroxylysine, and a C-terminal noncollagenous globular domain known as NC1 (25 kDa, 52 nm). The NC1 and 7S domains are key to the formation of the collagen network, while the triple helical domain is important in the integrity and flexibility of type IV collagen [14]. Using immunohistochemistry, collagen IV has been shown to remodel in a time-dependent manner during wound healing [162]. Collagen IV is also involved in subepidermal blistering diseases [163, 164].
1.2.3 Perlecan
Perlecan, often described as “pearls on a string” [15], is a 400–450 kDa heparan sulfate proteoglycan with five domains (I–V). The protein assembles into oligomers forming glycosaminoglycan (GAG) chains [16], which bind to type IV collagen’s NC1 domain and laminin α[alpha] chain G5 domain, while the core portion of perlecan binds to nidogen’s G2 domain and type IV collagen’s triple helix.
Studies have found that perlecans are the key organizing proteins in the BMZ, linking other important proteins, including type IV collagen and laminins [154]. Both the core portion and the GAG chains of perlecan regulate migration and proliferation by binding growth factors, including fibroblast growth factors, bone morphogenic proteins, platelet-derived growth factors, vascular endothelial growth factors, transforming growth factors, epithelial growth factors, hepatocyte growth factors, and insulin-like growth factor [17]. Perlecan is essential in angiogenesis through VEGF signaling [168], as well as neurogenesis, chondrogenesis, and the development of a variety of other tissues [17–21]. Increased perlecan expression has been reported in variety of odontogenic tumors [165]. On the other hand, perlecan-deficient mouse models report impaired corneal epithelium [166], as well as dysfunctional Schwann cells and peripheral nerve activity [167]. Furthermore, administration of domain V of perlecan was shown to be neuroprotective after ischemic strokes in mouse models, as well as angiogenic through increased VEGF expression [169].
1.2.4 The Laminin Family of Proteins
The most abundant BMZ noncollagenous proteins are the laminins. Eleven chains (five alpha, four β[beta], and three γ[gamma] chain), each with its unique gene, assemble into more than 16 different isoforms [23], each with a characteristic role in tissue development, structure, and function [24]. The α[alpha] chains have five homologous 150–180-amino-acid globular domains, together known as the G domain, not found on β[beta] and γ[gamma] chains and important in adhesion. The C-termini of the α[alpha], β[beta], and γ[gamma] chains interact to form a coiled-coil heterotrimer which gives rise to three short arms containing the N-terminus of each of the chains. The alpha subunits are critical for establishment of the BMZ network, attaching to various cell surface receptors, including integrins α[alpha]3β[beta]1, α[alpha]6β[beta]1, and α[alpha]6β[beta]4 [22, 23].
Laminin-111 and laminin-511 are large laminin isoforms that are critical during embryogenesis and have large N-terminal alpha chains which allow the proteins to self-polymerize. Laminin-511 is found in most BMZ, including the skin, kidney, and lungs, and important in ectoderm-derived tissues [22, 25]. Laminin-511 is produced by endothelial cells and important in angiogenesis [30], as well as wound healing by promoting proliferation and migration [25]. The protein is composed of a γ[gamma]1 chain, a β[beta]1 chain [26, 27], and an α[alpha]5 chain, which contains a G domain that binds to α[alpha]-dystroglycan and α[alpha]3β[beta]1 integrins [28, 29]. Furthermore, α[alpha]5-derived peptides may regulate metastatic activity and progression of breast cancer by interacting with cell surface proteoglycans [170, 172]. Laminin-511 as well as laminin-binding integrins are both critical in the maintenance of human embryonic stem cell pluripotency [171, 173].
1.3 Specialized BMZ: Intracellular Components
1.3.1 Plectin
Plectin, located in the innermost area of the hemidesmosome, is a 200 nm protein that links keratin intermediate filaments to the transmembrane collagen XVII [32] and α[alpha]6β[beta]4 integrin [33, 34] of the hemidesmosome [31]. The α[alpha]6β[beta]4 interaction with plectin is important in the formation and dissolution of the hemidesmosome [35, 36].
Clinically, patients with epidermolysis bullosa simplex with associated muscular dystrophy often are deficient in plectin resulting in an intraepidermal separation at the superior region of the hemidesmosome [37–40]. Epidermolysis bullosa simplex with severe mucus membrane involvement but without muscular weakness has been associated with nonsense mutations in plectin’s exon 32 [41]. Paraneoplastic pemphigus patients are reported to have plectin autoantigens [42]. Most recently, immunohistochemistry on head and neck squamous cell carcinoma revealed increased plectin expression was associated with the increased frequency of recurrence, poorer prognosis, and lower survivor rate, possibly through the MAP kinase pathway [174]. Plectin is important in tissue integrity and cell migration, and removal of plectin through siRNA impaired the attachment, migration, and invasion of a subtype of colon cancer cells [175]. Plectin-1, in addition, has been proposed as a potential biomarker for pancreatic ductal adenocarcinoma [176].
1.3.2 230 kDa Bullous Pemphigoid Antigen (BPAG1/BP230)
BPAG1 or BP230 is a 230 kDa protein located inside the cell. The protein has an N-terminal plakin domain, which binds to the ERBIN protein, and two homologous repeat domains on the C-terminus that bind to keratin filaments. BPAG1 has not been shown to be critical for the assembly of the BMZ or hemidesmosome [43–47].
Antibodies to BPAG1 are seen in the subepithelial blistering disease known as bullous pemphigoid [48–50]. In fact, an enzyme-linked immunosorbent assay (ELISA) to BP230 is available and helpful in the diagnosis of bullous pemphigoid [178, 179]. Antibodies to BPAG1 may also develop in conjunction to neoplasms leading to another blistering disease known as paraneoplastic pemphigus [51], as well as in patients with systemic lupus erythematosus [42]. A homozygous nonsense mutation in the gene coding for epithelial BPAG1’s coiled-coil domain has been identified in an autosomal recessive form of epidermolysis bullosa simplex [52]. Most recently, IgE autoantibodies against the C-terminus of BPAG1 were identified in a subtype of patients with pruritic dermatoses [177]. Anti-BPAG1 antibodies, furthermore, have been proposed as a serum marker for human melanoma [180].
1.4 Specialized BMZ: Transmembrane Components
1.4.1 Type XVII Collagen (BP180, 180 kDa Bullous Pemphigoid Antigen, BPAG2)
Type XVII collagen, also known as BPAG2 or BP180, is a transmembrane collagenous protein with a large C-terminus containing (as shown in Fig. 1.3) 15 interrupted collagen domains [53, 54]. The protein is important in adhesion and migration, possibly by connecting with laminin-332 [62, 181].
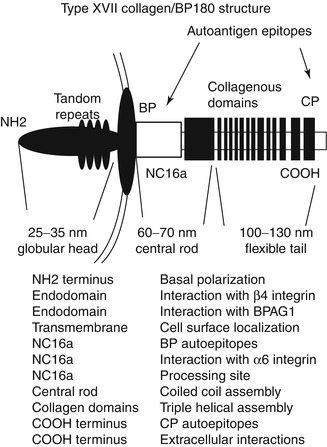
Fig. 1.3
Type XVII collagen/BP180. Type XVII collagen contains an intracellular globular head which interacts with β[beta]4 integrin and BPAG1/BP230. Extracellularly, the large noncollagenous NC16A domain contains the immunodominant bullous pemphigoid (BP) epitopes as well as sites for interaction with α[alpha]6 integrin and extracellular processing. Extracellularly, there is a central rod and a tail, which may allow come flexibility of the molecule. The collagenous domains are hypothesized to contain major linear IgA autoantibody epitopes, and the C-terminus contains epitopes recognized by cicatricial pemphigoid (CP) autoantibodies
Most of the protein is extracellular, is cleaved by ADAMS proteinases, and is shed from the cell surface [55, 56]. The intracellular portion of the protein binds to the hemidesmosome’s β[beta]4 integrin [61]. Type XVII collagen exodomain consists of an NC16A domain which lies adjacent to the largest collagenous domain 15 [59]. IgG autoantibodies develop to the noncollagenous NC16A domain which is important in some forms of bullous pemphigoid [57], and autoantibodies to this domain as detected by ELISA may be used as an indicator of disease activity [182]. In addition, autoantibodies develop to the distal portion of the central rod and flexible tail region in cicatricial pemphigoid [58], as well as lichen planus pemphigoides, localized vulval pemphigoid of childhood, and pemphigoid gestationis [60]. IgA autoantibodies target the extracellular portion of type XVII collagen in chronic bullous disease of childhood and linear IgA disease [60]. In some patients with generalized atrophic benign EB (GABEB), a nonlethal JEB, the absence of type XVII collagen has been found [63–65]. In fact, in a GABEB human xenograft model, correction of the condition was successful using ex vivo retroviral gene delivery of human collagen XVII cDNA [66].
1.4.2 Integrins of the Basement Membrane
Integrins are a group of heterodimeric receptor proteins that consist of α[alpha] and β[beta] subunits, each with small intracellular domains with the exception of integrin β[beta]4 which has a large 1,000-amino-acid intracellular domain. In the basal keratinocyte layer, two main integrins exist: α[alpha]6β[beta]4, found in the hemidesmosome [67], and α[alpha]3β[beta]1, found in the basal and lateral epidermis [68].
α[alpha]6β[beta]4 (Figs. 1.1, 1.2, 1.3, and 1.4) binds a variety of BMZ proteins including type XVII collagen and plectin. It also links extracellular laminin-332 with intracellular keratin filaments and plays a role in keratinocyte adhesion, migration, and proliferation [69, 70] through the NF-kβ[beta], mitogen-activated protein kinase-dependent pathway, or GTPase Rac1 pathway [71, 72]. The serine phosphorylation of the β[beta]4 subunit can lead to the dissolution of the hemidesmosome [48], while folding of the cytoplasmic portion of the protein is involved in the assembly of the hemidesmosome [73, 74]. Deficiency of β[beta]4 integrin can lead to significant blistering, dysfunctional adhesion, and the absence of hemidesmosomes in mouse models [75]. Mutant forms of α[alpha]6β[beta]4 are associated with epidermolysis bullosa with pyloric atresia (EB-PA) in which the epidermis separates above the plasma membrane with some portions attached to the dermis leading to mucocutaneous fragility and usually death in infancy [76–79]. In studies with transgenic mice, α[alpha]6β[beta]4 appears to help create a signaling environment that supports tumor development [183]. In addition, preliminary studies reveal that α[alpha]6β[beta]4 may be involved in regulation of colorectal cancer, as well as normal intestinal tissue [184]. Β[beta]4 in particular has been identified in its role in invasion of breast carcinoma through the SPARC (secreted protein acidic and rich in cysteine) protein [185]. While normal tissues lack α[alpha]3 G45 domain after proteolytic processing, SCC and wound edges show higher expression of this domain which binds to syndecan-1 [82, 89].
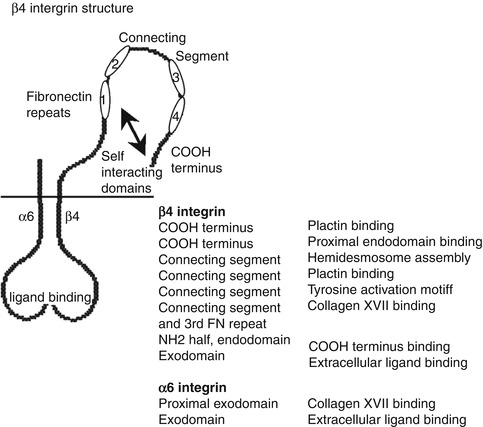
Fig. 1.4
Structure of β[beta]4 integrin. The especially large, approximately 1,000-amino-acid-long endodomain of β[beta]4 integrin contains four fibronectin-like repeats and a connecting segment separating repeats 1 and 2 from repeats 3 and 4. The endodomain has specific binding sites for plectin, type XVII collagen, and also has the ability to fold onto itself. Tyrosine and serine phosphorylation sites on the endodomain are thought to regulate hemidesmosome assembly, wound healing, and cancer development
1.5 Specialized BMZ: Extracellular Components
1.5.1 Laminin-332 and Laminin-311
Secreted by keratinocytes, laminin-332 is an extracellular protein with α[alpha]3, β[beta]2, and γ[gamma]2 chains (Fig. 1.5). Using the G domain of its α[alpha]3 chain, it binds to transmembrane α[alpha]6β[beta]4 integrins [80]. In addition, using its NC1 domain, it binds to type VII collagen [81]. After secretion, laminin-332 undergoes proteolytic processing [82] at the γ[gamma]2 chain and at two sites in the α[alpha]3 chain with C-proteinases like mammalian Tolloid and BMP-1 [83, 84], while the α[alpha]3 G domain cleavage site has also been shown to be acted upon by non-C-proteinases [85]. This proteolytic cleavage is important in cell growth and proliferation [86–88]. In fact, laminin-332 expression in SCC is associated with tumor invasion and prognosis [87]. While normal tissues lack α[alpha]3 G45 domain after proteolytic processing, SCC and wound edges show higher expression of this domain which binds to syndecan-1 [82, 89], which in turn promotes migration and invasion of tumor cells [88].
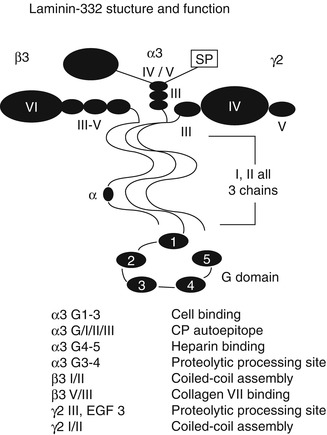
Fig. 1.5
Laminin-332 structure. Laminin-332 is a heterotrimer consisting of α[alpha]3, β[beta]3, and γ[gamma]2 subunits. The α[alpha]3 and γ[gamma]2 chains undergo proteolytic processing following secretion. Domains I and II of each chain interact together in trimeric assembly. The large C-terminal globular (G) domain on all laminin alpha chains, including the α[alpha]3 chain, contains 5 EGF repeats termed G1–G5 which bind cell surface integrin receptors as well as heparan sulfate-based receptors such as syndecan-1. The N-terminal laminin β[beta]3 domains III–V interact with the type VII collagen NC1 domain
By binding to different integrins, laminin-332 can regulate the adhesion of keratinocytes in vitro. Laminin-332 forms focal adhesions for attachment and migration by interacting with α[alpha]3β[beta]1 and can develop stable anchoring contacts by binding to α[alpha]6β[beta]4 [68, 90]. Focal adhesions can also evolve into anchoring contacts over time [35, 69]. Laminin-332 can also bind to NC1 domain of collagen VII using its β[beta]3 chain [81, 91].
In the lethal disease Herlitz junctional epidermolysis bullosa, there is an absence of laminin-332 [96, 97], while in nonlethal junctional epidermolysis bullosa, mutant laminin-332 is secreted but shows partial loss of function [93–95]. This decrease in laminin-332 function can cause substantial but nonlethal mucocutaneous blistering [99, 100]. Deletion of laminin α[alpha]3 domain IIIa can lead to mucocutaneous dysfunction known as laryngo-onycho-cutaneous (LOC) syndrome [102]. However, with replacement of laminin β[beta]3, phenotypical reversal can occur in keratinocytes lacking the β[beta]3 domain [98]. Furthermore, using a retrovirus ex vivo, normal laminin-332 β[beta]3 chain expression was successfully corrected with phenotypic improvement in a patient with non-Herlitz JEB [101]. Most recently, laminin-332 was identified as a pro-invasive protein in breast carcinoma [186]. Laminin-332 also controls sensory axon branching behavior and suppresses mechanotransduction, which could possibly provide yet another reason why some patients with laminin-332-deficient epidermolysis bullosa experience severe pain [187].
When the α[alpha]3 chain of laminin-332 combines with laminin γ[gamma]1 and β[beta]1 chains, another isoform of laminin is produced [82]. Known as laminin-311, it forms a disulfide bond with laminin-332 in tissue [103]; interacts with perlecan, dystroglycan, and integrins; and is important in signal transduction after mechanical stimulation [104].
1.5.2 Type VII Collagen
Found under the lamina densa, 426 nm type VII collagen is a crucial main constituent of the anchoring fibrils. The protein has a central collagenous region that is interrupted by noncollagenous domains. The noncollagenous regions, one of which is termed the hinge region, allow for type VII collagen to loop between interstitial collagen fibrils in the dermis with flexibility [105, 106]. Using ascorbic acid as a cofactor, the prolines on type VII collagen monomers are hydroxylated for additional stability [107]. These monomers assemble into homotrimer triple helices, and after C-proteinase proteolysis, the two trimers develop into antiparallel dimers [108, 109] and eventually into fully mature anchoring fibrils [110, 111]. Type VII collagen also links the lamina densa to the papillary dermis by interacting with other members of the BMZ, including type I [113] and type IV collagen [114].
Using the fibronectin repeat region of its NC1 domain, type VII collagen binds to laminin-332 [112]. In fact, IgG against the NC1 domain leads to a condition known as epidermolysis bullosa acquisita [81, 91, 128]. Defects in type VII collagen cause separation below the lamina densa leading to both the dominant and recessive cases of dystrophic EB [115–118], characterized by trauma-induced blisters mostly in the superior portion of the dermis [125]. The dominant form is often a result of a glycine substitution in the central repeat region [117], while in the recessive form of DEB, premature stop codons often result in the complete absence of anchoring fibrils [126, 127]. In some studies, recombinant human type VII collagen protein injections may phenotypically correct dystrophic EB, prolonging survival in animal models [119, 120]. Gene delivery of type VII collagen has also had some success in animal models [121–124].
1.6 Functional Role of BMZ Proteins
1.6.1 Development
As previously discussed, basement membrane proteins are important in the assembly and development of the tissue. Nidogen, important in the establishment of the basal lamina, exists as two isoforms with similar structures and functions. One is often upregulated if the other form is not present, as seen in nidogen-1 knockout mice. While the absence of one does not physiologically disrupt the BMZ, the absence of both can lead to severe skeletal, cardiac, and pulmonary defects that can be lethal [129]. In addition, the absence of both nidogen-1 and nidogen-2 can lead to severely impaired basement membrane formation of capillaries, with reduced deposition of other BMZ proteins and thin fragile walls without a well-defined basement membrane [188]. Laminin knockouts are usually lethal or extremely severe. Deletion of laminin α[alpha]2 is associated with congenital muscular dystrophy [130], while loss of laminin α[alpha]5 produces severe neurological, pulmonary, cardiac, kidney, and muscular defects [131, 132]. The absence of laminin-511 can lead to hair loss perhaps through the interruption of the sonic hedgehog and noggin signaling pathway needed for follicular downgrowth, while adding laminin-511 to deficient tissue corrects hair loss [133, 134]. In addition, human recombinant laminin-511 is now being utilized to aid long-term self-renewal of human pluripotent stem cells [173]. The absence of α[alpha]3 integrin produces hair defects very similar to that which occurs when laminin-511 is absent [138]. The absence of β[beta]1 integrin in mice models leads to abnormal deposition of other BMZ proteins, including laminin-332, leading to decreased hair matrix proliferation, hair loss, and blister formation [135, 136]. Skin cells deficient in α[alpha]3β[beta]1 show poor adhesion to the BMZ [137]. Mutations in type IV collagen, either the α[alpha]3, α[alpha]4, or α[alpha]5 chains, lead to Alport’s syndrome with abnormal basement membranes leading to retinopathy, deafness, and renal failure. Autoantibodies against the α[alpha]3 chain of type IV collagen produce a rapidly progressive glomerulonephritis with pulmonary dysfunction known as Goodpasture’s syndrome [139, 164].
1.6.2 Cancer
BMZ proteins are also important in the development of cancer. Laminin expression correlates with tumor invasiveness and prognosis [140–148] and is even essential to human SCC tumorigenesis as seen in xenograft studies involving laminin-332 [149]. Inhibition of laminin-332 α[alpha]3 G45 domain through deletion or antibody utilization in xenograft models inhibited human SCC tumorigenesis without affecting normal epithelial tissues [88], suggesting a role as a targeted cancer therapy in the future. α[alpha]6β[beta]4 integrin is also important in cancer progression, perhaps through signal transduction using the Y1494 residue of β[beta]4 [150]. In fact, recent data has shown that α[alpha]6β[beta]4 is a master transcription and translation regulator of other integrin subunits and especially important in wound healing and cancer [189]. Inhibition of β[beta]4 integrin through deletion or antibody utilization in xenograft models was also shown to inhibit human SCC tumorigenesis [149]. In mouse models of breast carcinoma, deletion of the of β[beta]4 integrin endodomain halted tumor progression, perhaps by inhibiting its interaction with laminin-332 [151]. Deletion of NC1 domain of type VII collagen also inhibited human SCC tumorigenesis [112], possibly by disrupting its interaction with laminin-332 and preventing phosphoinositol-3-kinase activation which is important in cellular invasion and proliferation [92].
1.6.3 Wound Healing
Laminin-332 and integrins β[beta]4 and β[beta]1 are also important in wound healing (Fig. 1.6). New studies which analyzed irradiated and wounded skin revealed decreased deposition of laminin-332. Keratinocytes with ionizing radiation not only had inhibited deposition of laminin-332 but also had slower migration, leading to delayed wound healing of irradiated tissue [191]. α[alpha]3β[beta]1 interacts with keratin filaments and is a component of focal adhesions in migrating cells [137]. β[beta]4 integrin’s N-glycosylation plays a role in adhesion and motility of keratinocytes [190]. It interacts with laminin-332, type XVII collagen, and plectin to form stable anchoring contacts that form hemidesmosomes. Focal adhesions over time can transform into stable anchoring contacts with the aid of plectin and CD151 [35, 69, 152]. Using β[beta]1 integrin and activated small RhoA GTPase, keratinocytes migrate over interstitial collagen at the leading wound edge. However, in areas farther from the leading edge, where laminin-332 is present, keratinocytes instead use β[beta]4 integrin and GTPase Rac1 signaling to migrate in a particular direction [70, 72, 153]. Thus, the presence or absence of laminin-332, which is dependent on its location within the healing wound, dictates which integrin is activated and how the cell migrates.
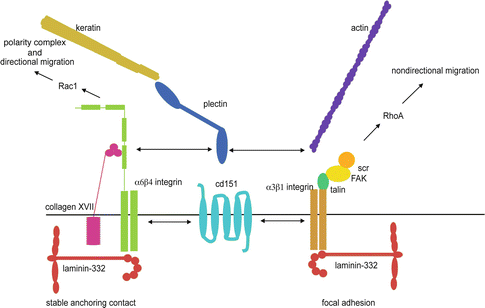
Fig. 1.6
Epithelial basement membrane components in wound healing and migration. Laminin-332 can interact with migratory cells via focal adhesions (FA) which contain α[alpha]3β[beta]1 integrin and supporting molecules including talin, FAK, and Src. Laminin-332 also localizes to complexes containing α[alpha]6β[beta]4 integrin, collagen XVII, and plectin, which are hemidesmosome-like structures termed stable anchoring complexes (SACs). While FAs associate with actin microfilaments, SACs instead associate with keratin intermediate filaments. Molecules such as CD151 and plectin can associate with both FAs and SACs and may help to facilitate the transition between these two structures. Engagement of β[beta]1 integrin with laminin-332 promotes RhoA GTPase activation, while binding of α[alpha]6β[beta]4 integrin to laminin-332 promotes GTPase Rac1 activation
References
1.
Timpl R, Dziadek M. Structure, development and molecular pathology of basement membranes. Int Rev Exp Path. 1986;29:1–112.PubMed
2.
Leblond CP, Inoue S. Structure, composition and assembly of basement membranes. Am J Anat. 1989;185:367–80.PubMed
3.
Keene DR, Marinkovich MP, Sakai LY. Immunodissection of the connective tissue matrix in human skin. Microsc Res Tech. 1997;38(4):394–406.PubMed
4.
Hippe-Sanwald S. Impact of freeze substitution on biological electron microscopy. Microsc Res Tech. 1993;24(5):400–22.PubMed
5.
Yurchenco PD, Smirnov S, Mathus T. Analysis of basement membrane self-assembly and cellular interactions with native and recombinant glycoproteins. Methods Cell Biol. 2002;69:111–44.PubMed
6.
Timpl R, Engel J. Type VI collagen. In: Mayne R, Burgeson RE, editors. Structure and function of collagen types. Orlando: Academic; 1987. p. 105–43.
7.
Kohfeldt E, Sasaki T, Gohring W, Timpl R. Nidogen-2: a new basement membrane protein with diverse binding properties. J Mol Biol. 1998;282(1):99–109.PubMed
8.
Miosge N, Kluge JG, Studzinski A, Zelent C, Bode C, Sprysch P, et al. In situ-RT-PCR and immunohistochemistry for the localisation of the mRNA of the alpha 3 chain of laminin and laminin-5 during human organogenesis. Anat Embryol (Berl). 2002;205(5–6):355–63.
9.
Mayer U, Nischt R, Poschl E, Mann K, Fukuda K, Gerl M, et al. A single EGF-like motif of laminin is responsible for high affinity nidogen binding. Embo J. 1993;12(5):1879–85.PubMedCentralPubMed
10.
Takagi J, Yang Y, Liu JH, Wang JH, Springer TA. Complex between nidogen and laminin fragments reveals a paradigmatic beta-propeller interface. Nature. 2003;424(6951):969–74.PubMed
11.
Ries A, Gohring W, Fox J, Timpl R, Sasaki T. Recombinant domains of mouse nidogen-1 and their binding to basement membrane proteins and monoclonal antibodies. Eur J Biochem. 2001;268(19):5119–28.PubMed
12.
Fleischmajer R, Schechter A, Bruns M, Perlish JS, Macdonald ED, Pan TC, et al. Skin fibroblasts are the only source of nidogen during early basal lamina formation in vitro. J Invest Dermatol. 1995;105(4):597–601.PubMed
13.
Marinkovich MP, Keene DR, Rimberg CS, Burgeson RE. Cellular origin of the dermal-epidermal basement membrane. Dev Dyn. 1993;197(4):255–67.PubMed
14.
Yurchenco PD, Furthmayr H. Self assembly of basement membrane collagen. Biogeosciences. 1984;23:1839–50.
16.
Iozzo RV. Basement membrane proteoglycans: from cellar to ceiling. Nat Rev Mol Cell Biol. 2005;6(8):646–56.PubMed
17.
Whitelock JM, Melrose J, Iozzo RV. Diverse cell signaling events modulated by perlecan. Biochemistry. 2008;47(43):11174–83.PubMedCentralPubMed
18.
Chuang CY, Lord MS, Melrose J, Rees MD, Knox SM, Freeman C, et al. Heparan sulfate-dependent signaling of fibroblast growth factor 18 by chondrocyte-derived perlecan. Biochemistry. 2010;49(26):5524–32.PubMedCentralPubMed
19.
Zoeller JJ, Whitelock JM, Iozzo RV. Perlecan regulates developmental angiogenesis by modulating the VEGF-VEGFR2 axis. Matrix Biol. 2009;28(5):284–91.PubMedCentralPubMed
Related posts:
 Kindlin-1 and Its Role in Kindler Syndrome
Kindlin-1 and Its Role in Kindler Syndrome
 Cyclophosphamide in Autoimmune Blistering Diseases: Safety, Efficacy and Evidence Base
Management of Bullous Systemic Lupus Erythematosus
Cyclophosphamide in Autoimmune Blistering Diseases: Safety, Efficacy and Evidence Base
Management of Bullous Systemic Lupus Erythematosus
 Using Intravenous Immunoglobulins in Autoimmune Bullous Diseases
Using Intravenous Immunoglobulins in Autoimmune Bullous Diseases
 Living with Epidermolysis Bullosa: Reviewing the Impact on Individuals’ Quality of Life
Living with Epidermolysis Bullosa: Reviewing the Impact on Individuals’ Quality of Life
 Dermatitis Herpetiformis
Dermatitis Herpetiformis
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


