!DOCTYPE html>
Stratum Corneum and Sensitive Skin
Stratum Corneum and Sensitive Skin
Subjects with sensitive skin report exaggerated reactions when their skin is in contact with cosmetics, soaps, and sunscreens, and they often report worsening after exposure to dry and cold climates. Epidemiologic studies have been carried out to assess whether there is a correlation with sex, age, skin type, or race and are described elsewhere in this book.
Subjects with sensitive skin may have a thinner SC with a reduced corneocyte area causing a higher transcutaneous penetration of water-soluble chemicals (1). Frosch and Kligman (2), by testing different irritants, showed a 14% incidence of sensitive skin in the normal population, likely correlated to a thin permeable SC, which makes these subjects more susceptible to chemical irritation.
Moreover, the declined barrier function in sensitive skin has already been reported as the result of an imbalance of intercellular lipid of SC (3). Although impaired barrier function is easily understood as a mechanism of sensitive skin, other factors are also possible implications such as changes in the nerve system and/or the structure of the epidermis. In a study (4), detailed characteristics of sensitive skin have been investigated using noninvasive methods. Sensitive skin has been classified into three different types based on their physiological parameters. Type 1 has been defined as the low barrier function group. Type 2 has been defined as the inflammation group with normal barrier function and inflammatory changes. Type 3 has been specified as the pseudohealthy group in terms of normal barrier function and no inflammatory changes. In all types, a high content of nerve growth factor has been observed in the SC, relative to that of nonsensitive skin. In both types 2 and 3, the sensitivity to electrical stimuli was high (4). Since these data suggest that the hypersensitive reaction of sensitive skin is closely related to nerve fibers innervating the epidermis, Yamasaki and Gallo (5) proposed that the innate immune system triggers an abnormal inflammatory reaction that mediates the symptoms of rosacea and sensitive skin. If so, flushing and blushing erythema may be due to chronic inflammation. In particular, cathelicidin may play a role in inducing the cytokine cascade. Indeed, some forms of cathelicidin peptides were known to have a unique capacity to be both vasoactive and proinflammatory (5).
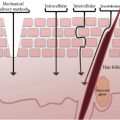
Direct connections were observed between unmyelinated nerve fibers and mast cells; stress in animal models induces substance P (SP) in unmyelinated nerve fibers, which triggers mast cell degranulation with subsequent histamine release (6). Stress is commonly reported as a trigger for sensitive skin, and mast cell degranulation is supported by the finding that sensitive skin sufferers had higher density of mast cells and size of lymphatic microvasculature (7). Neurogenic inflammation probably results from the release of neurotransmitters such as SP, calcitonin gene-related peptide (CGRP), and vasoactive intestinal peptide, which induce vasodilatation and mast cell degranulation. Nonspecific inflammation may also be associated with the release of interleukins. Indeed, sensitive skin could be the result of an inflammatory process resulting from the abnormal penetration in the skin of potentially irritating substances because of skin barrier dysfunction (8). In addition, the presence of a nonspecific reaction has been related to cutaneous sensory innervation in the establishment of skin sensitivity (9,10). Neuropeptides released from cutaneous nerves and skin resident cells such as SP, CGRP, and POMC peptides (such as β-endorphin and encephalin) are mandatory for a fine-tuned regulation of cutaneous immune responses and tissue maintenance and repair (11,12). In response to noxious stimuli, SP and CGRP lead to vasodilatation and mast cell degranulation, originating in a process called neurogenic inflammation. Classical pathways are then activated causing a nonspecific inflammation in consequence of released cytokines and eicosanoids such as interleukin-1 a (IL-1a), tumor necrosis factor alpha, prostaglandin E2, and prostaglandin F2 (13). On the other hand, POMC activities include antagonism and downregulation of adhesion molecules and reduced inflammation by the modulation of IL-10 production, which contributes to the amelioration of the subjective neurosensory forms of discomfort.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


