Fig. 18.1
Cutaneous leishmaniasis
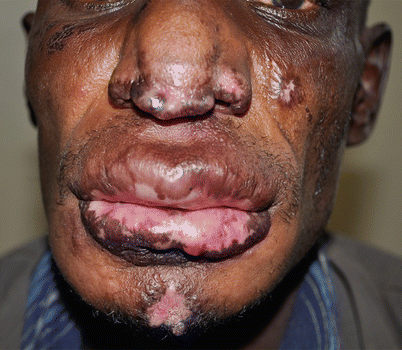
Fig. 18.2
Mucocutaneous leishmaniasis in an HIV-infected man
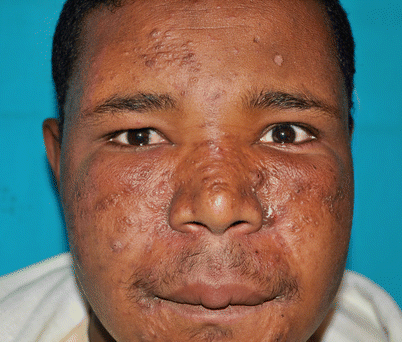
Fig. 18.3
Mucocutaneous leishmaniasis in an non-HIV-infected boy
Early Events in the Immune Response
In humans, the disease is initiated by the bite of an infected female sand fly (of the genus Phlebotomus in the Old World and Lutzomyia in the New World). During the bite parasites are inoculated into the dermis; in most cases less than 100 parasites are transmitted by a single bite, although the number may reach as high as 1000 or more [4].
Because the sand fly is a “pool feeder” (it severs capillaries in its hunt for a blood meal) promastigotes are deposited in a small pool of blood within the dermis. The bite of the sand fly initiates a local inflammatory response, with the recruitment of neutrophils, natural killer (NK) cells, eosinophils, and mast cells. The cellular response is rapid; neutrophils are the first cells to arrive at the site of the infection; in mice, they have been found collecting in the skin within 1 h of a parasite injection [5].
Most of the promastigotes deposited in the dermis are opsonized by serum complement and killed by complement-mediated lysis. The remaining promastigotes are phagocytosed via complement receptors expressed on the macrophage membrane, which binds to two abundant molecules on the surface of the parasites: gp63, a 63-kd neutral metalloproteinase; and LPG, a lipophosphoglycan [6].
Macrophages that have engulfed parasites release several chemokines, including CCL2, which attracts NK cells, dendritic cells (DCs), and additional macrophages to the site of the infection [7].
Components of sand fly saliva also have been shown to affect the immune response by exacerbating lesion development in resistant strains of mice [8, 9]. When maxadilan, a vasodilatory peptide isolated from sand fly saliva, was added to human peripheral blood mononuclear cells (PBMCs) the secretion of T-helper-1 (Th1) cytokines (interferon [IFN] and interleukin-12 [IL-12]) was decreased and the secretion of Th2 cytokines enhanced [10]. In addition salivary gland lysates (SGLs) also have been shown to augment the collection of inflammatory cells at the site of the bite and when added to experimental infections caused by L. major and L. braziliensis [11].
Once the promastigotes enter the macrophages, they transform into replicative amastigotes over a period of 2–5 days and are carried within modified lysosomal compartments known as parisitophorous vacuoles (PVs) [12]. A study of the ultrastructure of the vacuoles suggests a significant difference in packaging between Old World and New World leishmania. Amastigotes of L. major were found to be segregated into separate vacuoles during replication; in contrast, New World species of Leishmania were carried in large vacuoles occupied by many amastigotes [13].
Once inside the PVs the parasites produce antioxidant enzymes and are able to resist lysosomal hydrolases (the usual mechanism for clearing ingested pathogens). Within the cells the amastigotes replicate rapidly.
Macrophages that accumulate large numbers of replicating amastigotes rupture; the released parasites are taken up by neighboring competent cells. In especially inflamed skin lesions, most parasites are found in extracellular locations. One explanation is that high levels of inflammatory mediators interfere with receptors on the macrophages, blocking re-internalization of the parasites (Klaus S, unpublished data).
Antigen Transport and Presentation
At the same time that macrophages are ingesting parasites, Langerhans cells (LCs), which normally reside in the epidermis and are a potent type of antigen-presenting cell (APC), are stimulated. In a study of L. major infections in mice, LCs were found to be the dominant type of APC migrating from the skin to the regional lymph nodes via lymph channels [14, 15].
The ability of LCs (and other APCs) to identify the parasites has been attributed to Toll-like receptor 4 (TLR4), a member of a family of transmembrane receptors implicated in the recognition of a variety of microbial and foreign agents [16]. A study of TLR-deficient mice showed they had larger parasite burdens and were less efficient in the resolution of cutaneous lesions [17].
The extent to which macrophages also transport parasites and parasite antigen from the skin to the nodes is unresolved, although most studies indicate that LCs are the major carriers and shoulder most responsibility for antigen presentation to CD4+ naive T cells within the lymph nodes [18].
The initial stages of the uptake of antigen by the LCs are rapid; experiments in mice have demonstrated that LCs can find and engulf parasites within 4 h of exposure (although the migration to the lymph nodes may take up to 3 weeks) [19]. The movement by the LCs to the nodes is influenced by chemokine expression. Studies have indicated that during the migration the level of expression of chemokine receptor CCR7 on the LCs is enhanced, while the level of CCR2 and CCR5 is downregulated [20].
It is during this stage of the adaptive immune response that naive CD4+ T-helper cells (Th0) are programmed to develop into either Th1 or Th2 cells. Interleukin-12, a cytokine released by DCs, has a critical role at this stage in the immune response [21, 22]: IL-12, along with IL-1, serves as a promotor of the differentiation of Th1 cells in the lymph nodes from naive T0 cells. It also enhances chemokine gene expression in mice during the first 3 days of infection with L. major [23].
It is now clear that the successful clearing of amastigotes from infected macrophages in resistant mouse strains is mediated through the predominance of Th1 cells. The mechanism is through steps that link the production of IFN-γ, a major cytokine of Th1 cells, to the release of nitric oxide (NO), a compound within macrophages harboring amastigotes leading to their destruction. Nitric oxide ordinarily is present within an inactive form in the macrophages (iNO), which needs to be catalyzed by the enzyme NO synthase to become active. Interferon along with IL-12 and tumor necrosis factor (TNF), two other cytokines released by Th1 cells, make up the major upregulators of NO synthase, and the production of NO occurs only when the Th1 cells become predominant [24].
In contrast to the events within the resistant mice, in susceptible mouse strains (BALB/c) a type 2 response is initiated following infection with L. major, which tends to interfere with the protective activities. Th2 cells that become stimulated produce IL-4, IL-5, and IL-10, which have the capacity through mediators to inhibit the production of IFN-γ, downregulate the expression of iNOS (and consequently NO), and thus inhibit macrophage function.
Although a great deal is known about the process of resistance in mice, much less is known about the human immune response to Leishmania. While mice can mount either a Th1 or Th2 response, Rogers and Titus [25] suggested that in humans the principal response is predominantly, if not exclusively, a type 1 response. Using an in vitro system they cocultured PBMCs from Leishmania-naive donors with L. major parasites and found that type 1 cytokines were stimulated, (IFN-γ and IL12), and that when PBMCs were cocultured with macrophages infected with parasites, augmented intracellular killing was observed [25].
Antibody Response
Anti-Leishmania antibodies, as measured by enzyme-linked immunosorbent assay (ELISA), can be found in low titers in individuals recovering from CL, but their role in recovery from the acute infection and in the prevention of re-infection is debated. Polyclonal activation of human B cells leads to the production of large amounts of parasite specific antibodies, and amastigotes released into the dermis from ruptured macrophages appear to be coated with antiparasite antibodies [26].
In mice, antibody levels do not correlate in general with resistance to disease, although their effect on the course of the infection may depend on genetic factors. For example, passive transfer of antibody fractions from immune mice to BALB/c mice did not affect their susceptibility to infection [27], yet ablation of B cells in resistant mice generated a non-healing response to L. major [28].
Delayed-Type Hypersensitivity and the Leishmanin Skin Test
Delayed-type hypersensitivity in leishmaniasis can be measured by the leishmanin skin test (LST), in which the extent of a skin reaction is measured 48–72 h after an intradermal injection of 0.2 mL of a killed suspension of cultured leishmania promastigotes in saline. A positive reaction indicates a type 1 CD4+ cell-specific immune response. Because it is usually positive in individuals who have had CL, it has been used to measure the extent of infection among individuals living in endemic regions who have no history of overt disease. A study of 470 children living in endemic foci of L. major infection in Tunisia found that the proportion of asymptomatic infections among this group of children was approximately 10 % [29]. The LST also has been used as a predictor of susceptibility to subsequent disease, whether or not the individual had a history of a previous skin lesion.
Persistence of Parasites
In most cases of human cutaneous leishmaniasis it had been assumed that in skin lesions that had “healed” (either spontaneously or following treatment), the parasites had been eliminated by effector mechanisms involving IFN-γ and the generation of NO within the macrophages. It is now recognized that viable Leishmania organisms may persist in the skin long after the resolution of the clinical lesions [30]. Persistence has been documented in several ways: in biopsy samples from normal patients who have recovered from CL, in skin lesions of patients who recovered from CL but who later contracted HIV infection, and in skin lesions of patients who recovered from CL but later developed leishmania recidivans.
Schubach et al. [31] examined skin tissue obtained by biopsy at the sites of the scars from two patients from Brazil who had been infected with Leishmania 8 and 11 years earlier. The tissues from both patients grew out viable Leishmania parasites. A more recent study analyzed skin biopsies from scars of 32 patients with who had CL but who had been treated and clinically cured. Leishmania specific DNA was detected by polymerase chain reaction (PCR) in 30 of the patients, and parasites were isolated by culture in three [32].
The persistence of parasites in post-recovery CL is also evident in patients who later become infected with HIV. Studies of skin biopsies from such immunocompromised patients often show large numbers of amastigotes both within macrophages and free in the dermis. Parasites can also be seen within keratinocytes surrounding sweat ducts, and within the cells of the eccrine glands themselves [33]. Skin lesions in these patients often appear as isolated papules or plaques on exposed areas of the skin and are usually indistinguishable from similar lesions seen in non-immunosuppressed individuals. Skin lesions in HIV patients may also present as diffuse scaling plaques, which on biopsy show a high concentration of parasites [34].
Leishmaniasis recidivans (LR) is a rare clinical form of CL in which skin near the site of a previously healed acute CL lesion reappears as a dusky-red granulomatous plaque with active spreading borders. The clinical features resemble lupus vulgaris. Cultures of the skin lesion for Leishmania are usually negative, but with perseverance sparse parasites can be detected microscopically or by PCR. Leishmaniasis recidivans patients usually demonstrate high levels of antibodies in the serum and a strongly positive LST [35].
Vaccine Development
Clinical evidence points to the conclusion that recovery from skin infection with Leishmania provides lifelong protection against re-infection despite ongoing exposure to sand flies, suggesting that a vaccine would be of great value in controlling the disease.
For more than 100 years residents of endemic regions in eastern Asia and the Middle East practiced a form of vaccination known as leishmanization, which consisted of the deliberate inoculation of infective material into inconspicuous body areas (especially the buttocks) in the hope of providing protection from a subsequent infection and disfiguring scars on exposed parts of the body. A widescale trial of leishmanization was carried out among soldiers in the Iranian army in the 1990s, with more than a million individuals vaccinated, in this case using live L. major promastigotes obtained from cultures. Although the degree of protection initially seemed adequate (less than 3 % of a cohort of the vaccinated group developed a naturally acquired infection, compared with 14 % of unvaccinated volunteers), there was an unacceptable rate of adverse events: 2–3 % of the subjects developed large nonhealing infections at the site of the vaccinations that required treatment [36–38].
Today vaccine trials use killed or live attenuated parasites, genetically modified cells from promastigotes, and DNA encoding recombinant proteins [39–41].
Other vaccine candidates have included those using specific peptides derived from leishmania proteins such as amino acids derived from gp63, administered with certain adjuvants (such as liposomes or complete Freund’s adjuvant) [42]. One novel idea was that if components of sand fly saliva were added to a standard mix of antigens, a more effective prophylactic vaccine might result [43].
Although experimental vaccines to control CL have been studied extensively over the past two decades, inoculation with live L. major still remains the only successful vaccine in humans [44]. Currently, vaccine development is hampered by an incomplete understanding of the immune process and by concerns about long-term safety.
Onchocerciasis
Onchocerciasis, also known as river blindness, is a neglected tropical infection caused by the filarial nematode Onchocerca volvulus, with significant cutaneous manifestations, including pigmentary changes (Fig. 18.4), debilitating itching and subcutaneous nodules. It also causes significant eye damage and is the second (only to trachoma) most common infectious cause of blindness globally. It affects more than 18 million people living in endemic foci mainly of sub-Saharan Africa, but also in Latin America (including Brazil, Ecuador, Guatemala, Mexico, and Venezuela), and Yemen. It is now believed that skin disease is the most important contributor to the burden of onchocerciasis, rather than eye disease [45].
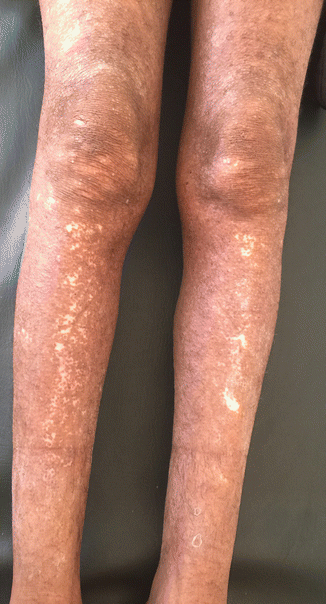
Fig. 18.4
Skin depigmentation in a patient with onchocerciasis
The disease is spread by the bite of black flies of the genus Simulium that prefer to breed on fast-running (highly oxygenated) rivers and streams of tropical countries. The female blackflies that transmit the disease need to feed on human blood for ovulation. Feeding on infected individuals results in ingestion of microfilariae of the parasite which then develop into infective larvae, also called third-stage filarial larvae (L3 larvae), inside the blackfly over the course of 2 weeks. Infection of humans occurs when these blackflies deposit L3 larvae while biting for another blood meal. Bites occur during the day time and multiple bites are required for disease transmission. Humans appear to be the primary host, although the gorilla in the Congo and the spider monkey in Mexico may also be naturally infected [46].
Once in the human host, the larvae migrate in the subcutaneous tissue forming nodules and slowly develop into adult worms over a course of about a year. Adult worms may live for 10–15 years and are usually palpable as firm, nontender nodules especially over the bony prominences of the pelvis, or on the scalp. From these sites, after a prepatent period of 3–18 months, fertilized female worms produce thousands of microfilariae daily (millions during a lifetime). The microfilariae (mfs), which can persist for 6–36 months, migrate to the subcutaneous and ocular tissues usually without provoking symptoms. In the dermis they are accessible to re-ingestion by blackflies, restarting the cycle. In addition to the skin, mfs can also be detected in the blood, urine and sputum. Most of the skin signs and symptoms of onchocerciasis, including severe pruritus and eye damage, are related to the body’s inflammatory response to dying and degenerating mfs.
The clinical features that characterize the disease include itching, commonly occurring over the lower trunk and buttocks, and an eczema-like eruption, which includes lichenification and hyperpigmentation, often called “lizard’s skin”. Individuals who have had the disease for many years may develop lymphedema and postinflammatory depigmentation, often on the anterior tibial surfaces (a sign called “leopard skin”). The prominent change seen on biopsy of affected skin is dermal fibrosis. In time, destruction of elastic tissue occurs, mediated by proteases from the parasites. These changes eventually lead to marked skin atrophy with redundant folds of skin in the inguinal areas, the so-called hanging groin. Two clinical patterns of reaction are evoked once the process is underway: a generalized form, characterized by widespread itching and dermatitis, and a hyperactive form, in which the skin reaction is often intense yet usually localized. The term “sowda” is reserved for severe hyperactive form that dominates one limb. A resistant form of the disease is also found among individuals living in hyperendemic areas and who remain unaffected despite being chronically exposed to the bites of infected flies. Decreased visual acuity is the most serious complication of the disease. The inflammatory reactions within the eye lead to iridocyclitis, choroiditis, and eventually optic atrophy.
The Role of Wolbachia Bacteria
The endosymbiotic Wolbachia bacteria, living inside both adult worms and mfs of O. volvulus, are essential for growth, development, fertility and survival of the parasite [47]. They are now considered to be an important target of the host’s inflammatory response [48, 49]. Neutrophil recruitment around the encysted adult worms (onchocercomas) appears to be related to the presence of the bacteria. In patients treated with doxycycline to eliminate the symbionts, the accumulation of neutrophils adjacent to the adult worms was drastically reduced [50]. Antibiotic treatment with doxycycline has also been found to improve skin lesions in hyperergic forms of the disease, and to interrupt embryogenesis of the female adult worms [51].
Immune Responses
Although early in the course of the disease—in the prepatent phase—inflammatory cells react to protein on the surface of the adult worms, the characteristic cutaneous signs and symptoms of the disease (dermatitis and itching) do not develop until mfs are produced by the gravid female worms. The mfs represent an ongoing source of antigen; up to 3000 are released daily by the adult female worm, beginning about 6–10 months after infection [52]. It is now argued that the predominant portion of the skin reaction in onchocerciasis is a reaction not only to the death and degeneration of the mfs but to their accompanying Wolbachia as well.
The intensity and type of host immune response gives rise to diverse clinical manifestations with two polar forms. The first, generalized onchocerciasis, occurs in hyporesponsive individuals having palpable nodules under their skin but no strong pathology despite carrying high mf skin loads, and the second, hyperactive form, is when patients exhibit severe inflammatory response with few worms [53].
Early in the course of the infection a polyclonal B-cell activation occurs with the production of parasite-specific immunoglobulins, including IgM, IgG (both IgG1 and IgG4), and IgE. The reaction to the mfs in the dermis and subcutaneous tissue is initiated by antibodies that attach to the surface of the parasites, along with complement. The immune complexes that are formed attract a variety of inflammatory cells, including neutrophils, eosinophils, and later macrophages. Degranulation of eosinophils appears to play the major role in the death of the mfs, but it is likely that proteases secreted by the larvae also add to the tissue damage. In addition to the effect on B cells, parasite antigens also induce substantial reactions from PBMCs. The initial recognition of these antigens, both the infective larvae themselves and their Wolbachia cargo, is mediated by TLRs. TLR4 responds to both the larvae of O. volvulus as well as to surface protein of the Wolbachia, and initiates a Th2 immune response mediated by IL-4 and IL-5 [54]. In addition, TLR2 also responds to surface protein isolated from Wolbachia, which mediates the release of TNF-α, IL-12, and IL-8 from PBMCs [55]. The evolution and final expression of the type of immune reaction that develops (Th1 or Th2) has been attributed to the early presence of specific cytokines; for example, the initial presence of IL12 directs the immune response toward a Th1 reaction, while a rapid induction of IL-4 promotes the generation of a dominant Th2-type response [56].
In some individuals an activation of a subset of CD4+ cells known as T regulatory cells (Tr1) occurs, which produce IL-10 and transforming growth factor (TGF), and some IFN [57].
In the usual generalized, chronic form of the disease, where the microfilarial load in the skin is high (up to 500 microfilaria per milligram of skin), the cutaneous reactions tend to be mild to moderate. In this type of onchocerciasis the cellular reactions are downregulated, with a suppression of Th1 and only a moderate Th2 response [58]. High levels of IL-10 are produced by CD4+ T-regulatory cells (Tr1), which act to inhibit activation of APCs and thus suppress proinflammatory functions. This type of immune response is thought to protect the host from acute skin damage (yet it may also be of benefit to the parasites by protecting them from some of the host’s lethal immune responses) [59]. In hyperreactive forms of the disease where the concentration of mfs is low (less than 10 per mg of skin) and the inflammatory reaction of the skin is severe, a strong Th2 response is seen. It is suggested that, in this form of onchocerciasis, inflammatory cells (eosinophils, neutrophils, and macrophages) all combine to kill mfs, under the direction of T cells and APCs.
Up to now HIV infection has been reported to play only a minor role in onchocerciasis, with no significant association with HIV detected in a large case control study [60]. No differences were noted in the density of mfs [61], although antibody response to the parasite was decreased in HIV-infected individuals, and they tended to lose their reactivity to these antigens over time [62].
Mazzotti Reaction
First described in 1948, Mazzotti reaction is a set of adverse reactions following administration of diethylcarbamazine (DEC) in patients with onchocerciasis. It has also been reported with administration of ivermectin and other antihelminths [63]. It is clinically characterized by intensely pruritic rash with or without systemic symptoms. Severity of the reaction correlates with the degree of microfilarial infestation and could at times be life threatening. A complex mechanism involving eosinophil degranulation and release of other mediators following the death of mf is involved in the pathogenesis. Patch testing using DEC is sometimes used to make a diagnosis of onchocerciasis [64].
Vaccine Development
Efforts to conquer onchocerciasis have been directed largely through the control of vectors and the use of community directed mass drug administration. With increasing treat of drug-resistance O. volvulus, there are compelling reasons to believe that vaccines will be the main tools for disease control in the future. Protective immunity against O. volvulus has been demonstrated in cattle [65] and mice [66]. In one study, 15 recombinant O. volvulus antigens out of the 44 screened using the O. volvulus-mouse model were found to be protective [67]. Eight of these antigens were produced and tested under controlled conditions, and only three, namely Ov-103, Ov-RAL-2 and Ov-CPI-2M, were able to repeatedly induce protective immunity [68]. Immunization in mice using DNA encoding of selected parasite genes has also shown promise [69]. A new initiative called The Onchocerciasis Vaccine for Africa (TOVA) is currently pursuing development of an onchocerciasis vaccine and has proposed two potential target product profiles, preventive vaccine for children less than 5 years old and therapeutic vaccine for infected individuals, with a plan to perform proof of concept trial for efficacy (phase two trials) at least for one candidate by 2020 [70].
Schistosomiasis
Schistosomiasis, also called bilharzia, is another neglected tropical infection in which schistosomal cercaria that normally parasitizes birds and mammals, including humans, cause skin disease or establish infection in the veins of the urinary or gastrointestinal tract, and occasionally, other organs. More than 260 million people are affected worldwide mainly in the tropical and subtropical regions [71].
The disease is caused by blood flukes of the family Schistosomatidae. Exposure to the parasite results from fecal or urinary contamination of freshwater that contains the intermediate host snails and skin contact to the same water by the definite host. Many eggs remain in the mammalian host causing an inflammatory reaction that results in morbidity. Less than half are released through the urine or feces depending on the infecting species. Each egg then releases a free-living stage of the parasite called miracidium that infects the intermediate host snail. Inside the snail miracidia undergo asexual reproduction giving rise to cercaria which are then released back into the water. Contact with contaminated water by the definite host results in penetration of cercaria through the skin that develop to maturity inside the body. When entering non-compatible hosts, parasites do not reproduce and die at various intervals after infection causing allergic reactions only [72].
During penetration and travel into the human skin cercaria transform to schistosomula causing human cercarial dermatitis (CD) or swimmer’s itch, represented by a maculopapular skin eruption associated with intense itching that occurs within few hours to days of exposure. Debilitating pruritus and constitutionals symptoms like diarrhea and fever may also occur based on the intensity of infection. Non-human schistosomes like Trichbilharzia are known to induce more severe skin reactions than human schistosomes, i.e. Schistosoma [72].
Another distinct skin manifestation of schistosmiasis is seen in Katayama syndrome (KS) or acute schistosomiasis. It is the result of immune-complex mediated response to the immature forms of the parasite and is characterized by extensive urticaria with or without angioedema, wheeze, fever, malaise, hepatosplenomegaly and other gastrointestinal symptoms that starts 2–12 weeks after exposure [73].
Related posts:





Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


