Summary and Key Features
- •
Some topical antioxidants (especially vitamins C and E, ubiquinone [coenzyme Q], and genestein) are effective in preventing ultraviolet damage to the skin (including sunburn, tanning, photoaging, and skin cancer) and can reverse the appearance of wrinkles and the pigmentation of solar lentigos.
- •
The formulation of each specific topical antioxidant is of utmost importance to assure stability in a commercial product with absorption by the skin and maintenance of activity after absorption.
- •
Both α-lipoic acid and its metabolite dihydrolipoic acid might protect the skin from oxidative stress, with similarities and differences in their activities and mechanisms of action. Animal and human studies have produced varied results, some demonstrating potential for adverse reactions.
- •
Topical as well as oral ubiquinone (coenzyme Q 10 ) act as both an antioxidant and a bioenergetic generator to retard intrinsic and extrinsic aging of the skin and to accelerate healing.
- •
Genistein, an isoflavone isolated from soy, decreases carcinogenesis by inhibiting tyrosine protein kinases (TPKs). As a potent antioxidant, topical genistein protects against photoaging and inhibits UVB-induced skin cancer. As a phytoestrogen, genistein enhances collagen synthesis in both intrinsic aging and photoaging and promotes healing.
See , , and .
Introduction
Dermatologists today have the capability to prevent damage to normal skin and even retard the skin’s natural aging through the use of cosmeceuticals. Especially during the past decade, research has demonstrated the efficacy of many topical nutrients, particularly antioxidants—some not synthesized by humans and therefore essential (vitamins C and E), some self-synthesized (α-lipoic acid, ubiquinone), and some exogenous (genistein). The challenge is to make topical formulations that attain percutaneous absorption of active forms and that maintain antioxidant activity. Such cosmeceuticals could protect as well as reduce and reverse manifestations of aging skin.
α-Lipoic Acid
R-Alpha lipoic acid (αLA) is synthesized in the mitochondria of plants and animals, including humans. Natural αLA is covalently bound to proteins via lysine; thus only minimal free αLA enters the circulation after biosynthesis or eating αLA-rich food. The lipoamide is a required cofactor for two enzymes in the citric acid cycle. It is also essential for the formation of a cofactor required in nucleic acid synthesis and for the metabolism of branched chain amino acids.
With oral supplements of free αLA, unbound αLA is transported to tissues. Free αLA is rapidly metabolized by the liver so that the half-life in blood after absorption is only about 30 minutes, limiting the amount delivered. High tissue levels are short-lived, since most free αLA is rapidly reduced to dihydrolipoic acid (DHLA), as shown in Fig. 25.1 .
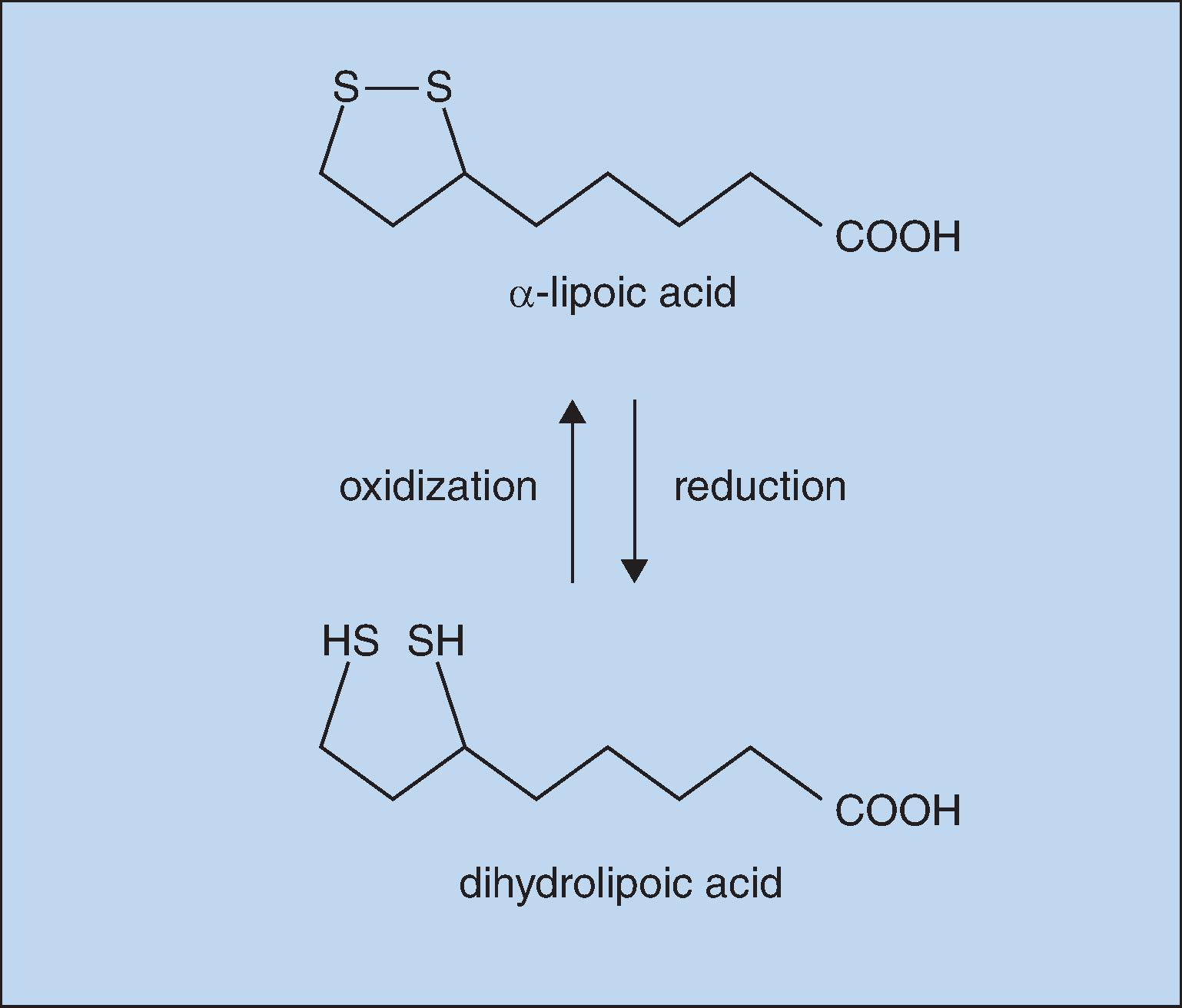
Notwithstanding this transient availability, free αLA has been shown to be therapeutic for autoimmune liver disease by binding autoantibodies, for heavy metal intoxication by trapping circulating metals, for diabetic polyneuropathy by preventing oxidative damage, and for mushroom poisoning. Although not normally found in significant amounts in the skin, αLA is a good candidate for topical application:
- •
As a small, stable molecule, it could successfully be percutaneously absorbed.
- •
As a potent antioxidant, it might protect from ultraviolet (UV) and other free radical environmental changes.
- •
Because it is soluble in both aqueous and lipid environments, it can interact with oxidants and antioxidants in many cellular compartments.
Indeed, αLA has been found to penetrate rapidly into murine and human skin to dermal and subcutaneous layers. Two hours after application of 5% αLA in propylene glycol, maximum levels of αLA were attained in the epidermis, dermis, and subcutaneous tissue. The stratum corneum concentration of αLA predicted the penetration and levels in the underlying skin. Five percent of the αLA was converted to DHLA in both the epidermis and dermis, leading the researchers to conclude that both keratinocytes and fibroblasts reduce αLA.
DHLA, unlike αLA, has the capacity to regenerate the endogenous antioxidants vitamin E, vitamin C, glutathione, and ubiquinol, as illustrated in Fig. 25.2 . This is clearly of great importance for skin, since UV exposure directly depletes ubiquinone (coenzyme Q 10 [CoQ 10 ]) and vitamin E in particular, as well as vitamin C, thereby stressing the other linked antioxidants. Regeneration of these major membrane and cytosol antioxidants gives cascading protection. Increases in the other important antioxidants (intracellular glutathione and extracellular cysteine) are noted when αLA is added to cell cultures. Vitamin E–deficient animals do not show symptoms (weight loss, neuromuscular abnormalities) when supplemented with αLA.
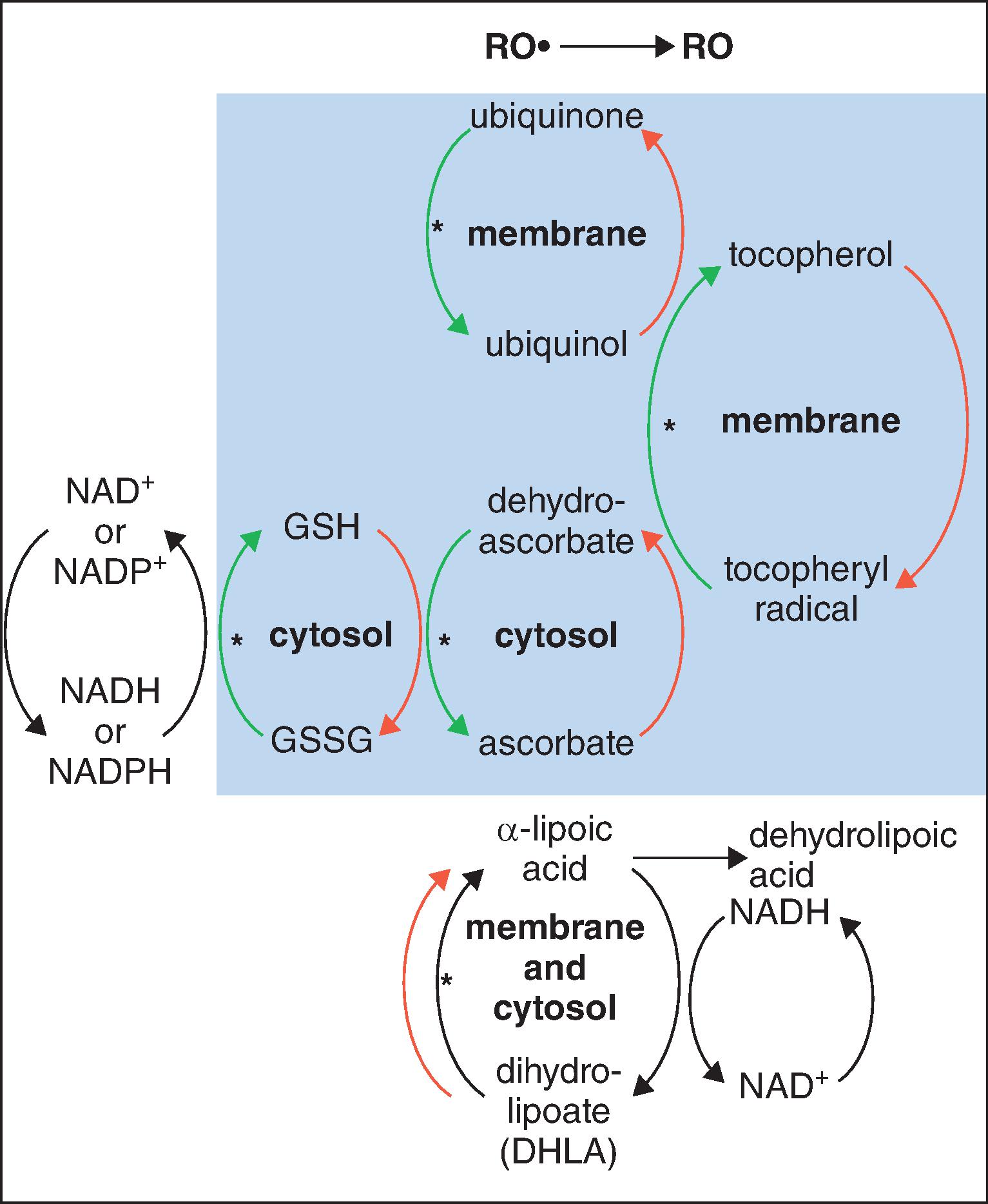
Topical αLA with its metabolite DHLA might directly protect the skin from oxidative stress in several ways. Both αLA and DHLA are highly effective antioxidants as summarized in Table 25.1 . DHLA is actually the more potent form. Both successfully scavenge reactive oxygen species (ROS) in vitro and in vivo . However, prooxidant activity has been observed. This occurs when an antioxidant reacts with an ROS scavenger, forming a product that is more harmful than the scavenged ROS. Fortunately, αLA can act as an antioxidant against the prooxidant activity of DHLA. Both αLA and DHLA further provide antioxidant activity by chelating Fe 2+ and Cu 2+ (αLA) and Cd 2+ (DHLA).
| α-Lipoic Acid | Dihydrolipoic Acid | |
|---|---|---|
| Antioxidant | + | ++ |
| Scavenges reactive oxygen species | + | + |
| Chelates metals: | ||
| Fe 2+ , Cu 2+ | + | – |
| Cd 2+ | – | + |
| Regenerates endogenous antioxidants (vitamin E, vitamin C, glutathione, ubiquinol) | – | + |
| Repairs oxidatively damaged proteins | – | + |
| Prooxidant | + | + |
Also, αLA (but not DHLA) acts as an antiinflammatory agent by reducing the production and inhibiting the binding of transcription factors such as nuclear factor-kappa beta (NF-κβ), thereby indirectly affecting the gene expression of inflammatory cytokines such as tumor necrosis factor-alpha (TNF-α) and interleukins (ILs). DHLA (but not αLA) can repair oxidatively damaged proteins, which in turn regulate the activity of proteinase inhibitors such as α 1 -AP, an inflammatory modulator. In vitro , both αLA and DHLA inhibit lipolysaccharide-induced nitric oxide (NO) and prostaglandin E 2 (PGE 2 ) formation and suppress inducible NO synthase (iNOS) but do not affect the expression of cyclooxygenase-2 (COX-2). In a mouse model, topical DHLA inhibits chemically induced activation of skin inflammation with a concomitant decrease in inflammatory modulators. Furthermore, topical DHLA (but not oral αLA) reduces chemically induced skin tumor incidence and multiplicity, and inhibits iNOS and COX-2 in a dose-dependent manner. As antioxidants, both αLA and DHLA are directly antiinflammatory by virtue of their quenching oxidants secreted by leukocytes and macrophages at sites of inflammation.
As a potential antioxidant with an antiinflammation activity, αLA might be an excellent topical cosmeceutical. Unfortunately this molecule is not stable because of its low melting point and the distorted dithiolane ring that absorbs light around 330 nm (low-wavelength UVA), causing decomposition. However, when αLA is exposed to UVB in the presence of thiol compounds such as cysteine and homocysteine, but not methionine, this decomposition of the αLA molecule is retarded and, instead, increased recovery of αLA ensues. Thus possibly formulating αLA with cysteine or homocysteine might improve its photochemical stability. The dithiolane ring structure further renders αLA quite vulnerable to thermal stimuli, causing decomposition and formation of interlocking polymers. This problem is more difficult to surmount when making a product with enough stability to assure an effective shelf-life.
Thus whether αLA can provide effective protection against UVB-induced damage is not clear. A single topical application of αLA (0.5 µmol/cm 2 ) on pig skin was shown to reduce UVB radiation–induced oxidative stress and lipid peroxidation, thereby reducing apoptosis. In contrast, other research reported that topical application of 5% α-lipoic acid is ineffective at suppressing UV-induced sunburn cells as evaluated by apoptotic markers in keratinocytes. Another study demonstrated that αLA did protect against ionizing radiation–induced lipid and protein oxidation in mice and significantly decreased the formation of malondialdehyde (MDA, a marker of oxidative stress) in all tissues examined, especially in the brain, most probably due to its free radical–quenching ability.
αLA enhances repair of age-induced damage to skin barrier function and to epidermal and dermal structural integrity, as demonstrated in vitro with a new “skin-on-a-chip” technology. Treatment of human skin equivalents incubated with 10µm αLA increased filaggrin gene expression and increased the synthesis of the epidermal structural proteins filaggrin, involucrin keratin 10, and collagen IV. These antiaging improvements in the epidermal layer were histologically noticeable.
αLA and DHLA have been shown to be effective depigmenting agents. Both depigment dark-skinned swine, inhibit tanning of light-skinned swine, and inhibit chemical and UVB-induced tyrosinase activity in melanocyte cultures. A recent new derivative of α-lipoic acid has been proven to be an effective depigmenting agent in melanoma cells in vitro . This depigmentation is achieved by formation of DHLA conjugate products.
αLA may prove to retard and correct both intrinsic and extrinsic aging of the skin as well as other organs. By damaging DNA, the ROS continuously formed in normal metabolism may be largely responsible for the functional deterioration of organs with aging. A decrease in cellular protein and DNA as well as in αLA levels has been measured in aged rat liver, kidney, and spleen. Supplementation with αLA increases nucleic acid and protein levels in the elderly organs. Similarly, the age-related decrease of mitochondrial function in cardiac and brain cells can be improved with αLA supplementation. Clearly, aging skin might similarly benefit.
Another symptom of aging is a loss of pressure-induced vasodilation (PIV). The interaction of primary afferent nerves with the skin’s vascular endothelium maintains PIV to preserve homeostasis of blood flow to the skin for optimal oxygenation. PIV is thus a cutaneous protective mechanism to prevent, for example, pressure ulcers. In aging, PIV is compromised due to endothelial dysfunction (probably from an alteration in the nitric oxide pathway). Aging and inflammation cause oxidative stress that leads to endothelial cell and nerve damage; also, noncontrolled hyperglycemia in diabetes causes oxidative stress that adversely affects PIV. αLA is able to restore PIV, possibly through reduction of oxidative stress.
To evaluate possible improvement of photodamage, a split-face study was done on 33 females. Topical application twice daily of 5% lipoic acid cream for 12 weeks decreased skin roughness by 50.8% (as measured by laser profilometry) when compared with the placebo. Clinical and photographic evaluation showed reduction in lentigines and fine wrinkles. In another study, twice-daily oral intake of αLA combined with other proteins, vitamins, and minerals improved wrinkles, roughness, and telangiectasias after 4–6 months in humans, as assessed clinically and by measurements of skin thickness and elasticity. Experiments with fibroblasts in vitro showed increased collagen synthesis when high concentrations of αLA were added to culture media.
Clearly, topical αLA should be further studied by quantitative techniques to confirm these results and to elucidate mechanisms of action. Whether αLA modulates signaling pathways by scavenging ROS as an antioxidant or by direct inhibition of a signal transduction like an enzyme is difficult to determine. The benefits of avoiding the UV-related skin inflammatory damage by αLA may be accompanied with risks such as induction of skin cancer. In fact, there are no reports of the inhibitory effect of LA on photocarcinogenesis. Careful application of αLA for skin aging must be recommended to avoid unwanted adverse effects. Further basic data on αLA are absolutely necessary to assure stability for long and safe topical use of αLA.
Ubiquinone (Coenzyme Q 10 )
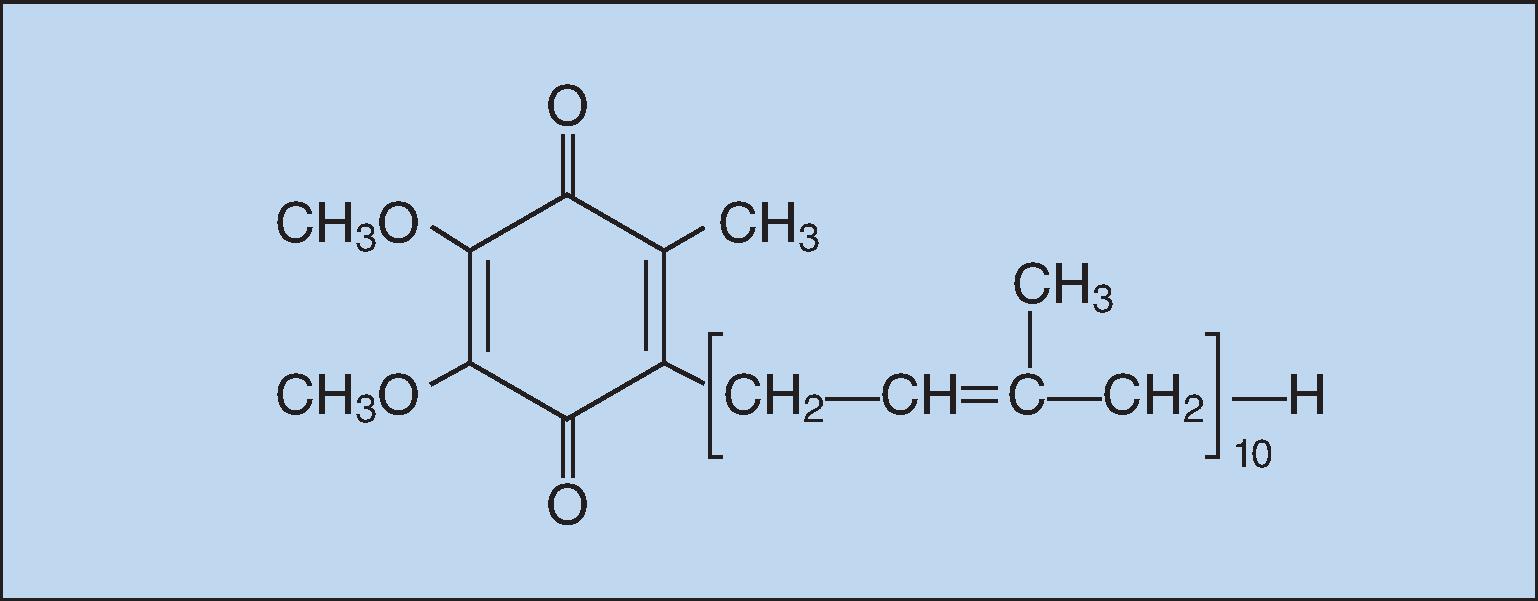
Ubiquinone (coenzyme Q 10 [CoQ 10 ], Fig. 25.3 ) is so named because it is ubiquitous in virtually all living cells, excluding some bacteria and fungi, although the level is quite variable. Since most human tissues synthesize CoQ 10 , it is not considered to be a vitamin.
CoQ 10 is primarily located in the inner mitochondrial membrane where it is essential for the production of the adenosine triphosphate (ATP) required for all vital cellular functions. Until recently, CoQ 10 was thought to function only in energy transduction: indeed, exogenous CoQ 10 has proven very important in ameliorating bioenergetic impairments not only in myopathies and cardiomyopathy, but also in aging skin. With the relatively recent discovery that CoQ 10 is also an antioxidant within subcellular membranes, new roles are now being recognized:
- 1
CoQ 10 can regenerate reduced tocopherol, as depicted in Fig. 25.2 . In fact, within membranes, the amount of CoQ 10 is from 3 to 30 times that of tocopherol. Without CoQ 10 , the regeneration of tocopherol would be very slow.
- 2
In the skin, CoQ 10 not only preserves antioxidant defense mechanisms that otherwise decrease with intrinsic aging but also enhances these defenses to protect against extrinsic (especially UV) damage.
- 3
Through gene induction, CoQ 10 increases the synthesis of collagen and elastin and decreases the expression of metalloproteinases that degrade collagen.
- 4
CoQ 10 inhibits UV activation of proinflammatory cytokines.
The concentration of CoQ 10 is highest in organs with high rates of metabolism such as heart, kidney, and liver, where it functions as an energy transfer molecule. In skin, the level of CoQ 10 is relatively low, with 10-fold higher levels in the epidermis than in the dermis. Thus the epidermis in particular might potentially benefit from topical CoQ 10 . It has been demonstrated that CoQ 10 can be topically absorbed: application of CoQ 10 in ethanol to porcine skin achieved 20% penetration into the epidermis and 27% into the dermis.
Though oral supplementation does not increase levels in the dermis or other organs, it does increase serum and epidermal CoQ 10 substantially. This increase is important because UV irradiation depletes CoQ 10 as well as other antioxidants in skin, enhancing oxidative damage. Sebum is among the first skin components to be directly confronted with exogenous oxidative insults. UV depletes 84% of vitamin E and 70% of CoQ 10 from sebum with a decrease of 30% of the lipid component squalene. With exposure in the absence of vitamin E and CoQ 10 , the same dose of UV produces a 90% decrease in squalene. Therefore vitamin E and CoQ 10 work synergistically to inhibit the UV-induced depletion of squalene and other unsaturated lipid molecules.
Environmental airborne pollutants from traffic, industry, cooking fuel, cigarette smoke, and ozone cause premature aging of the skin as well as skin precancers and cancers. Particulate matter (PM) pollutants activate dermal mast cells to degranulate, causing inflammatory insult to dermal tissue and fibroblasts, which generates extra- and intracellular ROS as well as mitochondrial ROS. In vitro studies demonstrated that CoQ 10 ameliorates this cellular oxidative damage and suppresses the damaging inflammatory cascade induced by PM.
The antiaging efficacy of CoQ 10 was further seen in an in vitro skin-on-a-chip model. As CoQ 10 concentration increased, the number of cells/cm 2 and thickness of the epidermal layer increased, with a dose-dependent increase in levels of filaggrin.
Interestingly, statin medications frequently prescribed to decrease hypercholesterol target an enzyme in the mevalonate pathway that is required for synthesis of cholesterol and isoprenylated compounds such as CoQ 10 . When the statin concentration is >10,000 nm, CoQ 10 deprivation follows with consequent intracellular oxidative stress, mitochondrial dysfunction, and death of dermal fibroblasts. At lower statin concentrations, cells (particularly mitochondria) can adapt to prevent overt toxicity. The natural decrease in CoQ 10 with intrinsic aging promotes increased mitochondrial permeability and bioenergetic dysfunction leading to premature aging of fibroblasts as demonstrated in vitro.
Thus CoQ 10 has the potential to inhibit intrinsic as well as extrinsic aging of the skin because it serves as an energy generator as well as an antioxidant and a regulator of gene induction. By maintaining a proper energy level in the mitochondria, CoQ 10 prevents aging cells from switching to anaerobic energy production. Cutaneous aging is characterized by a decline in the mitochondrial energy metabolism of skin cells, mediated by free radical reactions arising both endogenously (ROS generation by increased “leakage” of electrons from the respiratory chain) and exogenously (UV exposure). This impairment of mitochondrial function hinders cellular ATP synthesis, reducing the “fuel supply” for repair mechanisms. By-products of impaired mitochondrial respiration induce formation of ROS that, in turn, damage neighboring mitrochondrial complexes, membranes, and mitochondrial DNA (mtDNA) leading to mutations, thus accelerating the aging processes in a feedback loop. Furthermore, with loss of mitochondrial energetic capacity, there is a switch to anaerobic pathways such as glycolysis, with the formation of advanced glycation endproducts (AGEs) that harm cells by generating nonfunctional cytoskeletal proteins and inducing apoptosis.
CoQ 10 retards intrinsic aging of the skin, not only as an energy generator but also as an antioxidant, as confirmed in vitro by sophisticated ultraweak photon emission (UPE). Increased antioxidation results in a decreased UPE. Elderly volar skin demonstrated a 33% reduction in antioxidant activity when compared with young skin. This was corrected after 1 week of twice-daily topical application of 0.3% CoQ 10 .
In the skin, CoQ 10 also preserves intrinsic antioxidant defense mechanisms: with intrinsic aging, there is a detrimental increase in the activity of phosphotyrosine kinase, a loss of glutathione, and oxidative damage to DNA. All of these manifestations of intrinsic aging are prevented by treatment with CoQ 10 .
CoQ 10 has been proven to retard intrinsic aging of the skin by direct gene induction. Aged human fibroblasts in vitro proliferate more slowly than young cells and produce less collagen, elastin, and hyaluronic acid. The addition of CoQ 10 has previously been shown to increase rates of cell division and levels of glycosaminoglycan in neonatal fibroblasts. Recent experiments on primary embryonic, neonatal, adolescent, and adult human dermal fibroblasts confirmed that CoQ 10 treatment increases proliferation of fibroblasts and enhances synthesis of type IV collagen. This research further demonstrated an increase in elastin synthesis. The increase in collagen and elastin production parallels enhanced type IV (and type VII) collagen and elastin gene expression. Fibroblasts of all ages were equally responsive.
Because human fibroblasts in vitro provide an excellent, well-established experimental model for senescence, Japanese scientists, faced with an aging society, have pursued investigations of CoQ 10 for skin care. One recent study demonstrated that CoQ 10 treatment of human fibroblasts in vitro did indeed increase intracellular CoQ 10 concentrations and promote proliferation of fibroblasts as well as increased mRNA expression of type I, IV, and VII collagens, elastin, and heat shock protein 47 (HSP47, a collagen binding chaperone glycoprotein in the endoplasmic reticulum responsible for the maturation of collagen I and II molecules). Similarly when human fibroblasts in vitro were exposed to damaging hydrogen peroxide, CoQ 10 suppressed the oxidative stress and degradation of collagen and increased cell viability as well as mRNA levels of type I and IV collagen.
Protection from extrinsic UVA- and UVB-induced photoaging has been extensively demonstrated by many studies. In hairless mice, UVB irradiation led to a significant lowering of manganese superoxide dismutase (SOD 2 ) and glutathione peroxidase (GPx). The application of CoQ 10 to the UVB-irradiated skin significantly increased SOD 2 and GPx. Similarly, after UVA irradiation, a decrease in antioxidant activity was noted, but this loss was significantly corrected with topical 0.3% CoQ 10 . Further research showed that CoQ 10 (0.3%) also suppresses the UVA-induced reduction of mitochondrial membrane potential in keratinocytes and fibroblasts cultured from both young and old donors (with amelioration of 53%). Supplemental CoQ 10 was further effective against UVA-mediated oxidative stress in cultured human keratinocytes exposed to hydrogen peroxide.
Very importantly, CoQ 10 protects from the UVA-induced degradation of collagen. Both CoQ 10 and vitamin E were shown in vitro to suppress fibroblast production of UVA-induced collagenase, thereby markedly retarding collagen breakdown. CoQ 10 suppresses collagenase expression over a longer period of time than does vitamin E. The recent experiments cited earlier (demonstrating inhibition of intrinsic aging with in vitro fibroblasts of several ages) confirmed protection from extrinsic UV compromise of antioxidant function: treatment with CoQ 10 decreased UV-induced production of intracellular ROS and inhibited UV activation of matrix metalloproteinase-1 (MMP-1), thereby preventing UV-induced collagen breakdown. UV activation of preinflammatory cytokines, specifically Il-1α, was also inhibited, thus protecting from UV-induced inflammation and the resultant appearance of premature aging.
Other in vitro research provided evidence of photoprotection by CoQ 10 from UVA. Human dermal fibroblasts supplemented with a formulation of CoQ 10 (also containing a sun protection factor [SPF] 10 sunscreen with a strawberry extract) provided significant photoprotection, decreasing ROS, increasing antioxidant defense, decreasing fibroblast apoptosis, lowering inflammatory markers, and improving mitochondrial function.
Photoprotection was further demonstrated by exposure of mice to UVB. Treatment with CoQ 10 “sunscreen” prevented not only the UVB-induced decrease in antioxidant enzymes (SOD, GPx, and DNA [cytosine-5]-methyl transferase) but also the damaging increase in MDA activity and MMP-1.
The efficacy of CoQ 10 in reversing photoaging by preserving collagen and elastin was further studied clinically. CoQ 10 cream (0.3%) was applied to one-half of the face and placebo to the other, once daily for 6 months. Casts were made of the periorbital rhytides. Quantitative microtopography demonstrated a 27% reduction in the mean wrinkle depth.
Oral CoQ 10 was also effective in reversing the appearance of aging. A liquid food supplement containing water-soluble CoQ 10 with collagen improved dermal density and periorbital wrinkle area, total wrinkle score, and skin smoothness after 12 weeks of once-daily intakes. Unfortunately there were no significant changes in skin hydration, dermal thickness, transepidermal water loss (TEWL), or viscoelasticity.
Another clinical measure of photoaging is stratum corneum cell size. With deceased cell turnover time in aged skin, corneocytes become larger. Treatment once daily for 6 months with CoQ 10 cream decreased corneocyte size equivalent to rejuvenation of 20 years.
Hyperpigmentation is an unattractive consequence of photoaging that possibly can be effectively treated with CoQ 10 . B16 melanoma cells were exposed to CoQ 10 in vitro . Melanin synthesis was decreased in a dose-dependent manner with a corresponding dose-dependent decrease in tyrosinase activity. The highest concentration of CoQ 10 (2 μM) was as effective as ascorbic acid (5 mM) in inhibition.
Although CoQ 10 does decrease melanin synthesis in vitro in melanoma cells (and presumably also in normal melanocytes), overall CoQ 10 has the adverse effect of enhancing survival of these deadly cells. The enzyme UBIAD1 (UBIA prenyltransferase domain–containing protein 1), which biosynthesizes nonmitochondrial CoQ 10 , may be partially responsible for poor overall survival of melanoma patients. UBIAD1 and CoQ 10 levels are unregulated in melanoma cells as compared to normal melanocytes: the increased plasma membrane CoQ 10 can prevent lipid peroxidation that would otherwise lead to melanoma cell death.
A lower-molecular-weight, short-tail analog of CoQ 10 , idebenone, has also been shown clinically to repair photodamaged skin. In a nonvehicle controlled study, 41 females aged 30–65 years applied either 0.5% or 1.0% idebenone twice daily for 6 weeks. Use of 1.0% idebenone increased skin hydration by 37% (as measured by electrical conductance), reduced skin roughness/dryness by 26%, reduced fine wrinkles by 29%, and improved photoaged skin by 33% (as subjectively evaluated). The 0.5% idebenone was almost as effective as 1.0%. Immunofluorescence and staining of punch biopsies revealed a decrease in IL-1b, IL-6, and MMP-1 and an increase in collagen for both concentrations. However, idebenone did not improve cellular bioenergetics in CoQ 10 deficient fibroblasts in vitro .
A final advantageous action of CoQ 10 is acceleration of healing. Oral CoQ 10 , given pre- and post-treatment, was shown to enhance healing after laser resurfacing and chemical peels as measured by accelerating reepithelialization.
Thus CoQ 10 rejuvenates the skin by (1) enhancing mitochondrial energy generation, (2) increasing fibroblast cell division and synthesis of collagen and elastin, and (3) as an effective antioxidant, protecting the epidermis and the dermal matrix from both intrinsic and extrinsic aging. Recent extensive research continues to demonstrate new advantageous functions of CoQ 10 for the skin, making this potentially a very important, effective cosmeceutical.
Genistein
Genistein is an isoflavone cosmeceutical isolated from soy. Recent interest in genistein has been stimulated by epidemiologic studies that correlate diets high in soy with reduced incidence of cardiovascular disease, osteoporosis, and certain cancers in humans.
The direct anticarcinogenic action of genistein is documented. Animal studies demonstrate protection against bladder, breast, colon, liver, lung, prostate, and skin cancers with oral genistein, and dietary soy inhibits chemically induced skin cancer in mice. Growth of many in vitro cancer cell lines is inhibited by genistein. Genistein also arrests the growth and induces the differentiation of malignant melanoma cells in vitro and inhibits pulmonary metastases of malignant melanoma cells in vivo . Decreasing metastases may be through genistein’s proven reduction of neoplastic cell migration and inhibition of angiogenesis.
The mechanism by which genistein inhibits carcinogenesis may be through inhibition of tyrosine protein kinases (TPKs), the enzymes which phosphorylate proteins necessary for the regulation of cell division and transformation. Of particular importance is phosphorylation of TPK-dependent epidermal growth factor receptors (EGF-R), which are related to tumor promotion, including initiation of transcription factors, release of inflammatory mediators (as prostaglandins), and stimulation of cell proliferation. Genistein was found to downregulate both UVA- and UVB-induced EGF-R phosphorylation in human epidermoid carcinoma cells in vitro . By directly inhibiting activation of nuclear factor-kappa beta (NF-κβ), and therefore the resultant inflammatory cytokines, genistein further protects. In mouse skin, genistein also blocks the UVB-induced expression of the photooncogenes c- fos and c- jun , which promote cell proliferation in oncogenesis. Similarly, genistein retards UV-induced apoptotic changes—including caspase-3 and p21-activated kinase 2 activation of human epidermal carcinoma cells and phosphokinase Cδ in human keratinocytes.
Genistein is also a potent antioxidant. Genistein scavenges peroxyl free radicals, thereby protecting against lipid peroxidation in vitro and in vivo . The decreased incidence of cardiovascular disease with high-soy diets may be due to genistein’s inhibiting the oxidation of low-density lipoprotein (LDL) cholesterol in both aqueous and lipophilic environments. Elegant studies on human diploid fibroblasts (HDFs) further elucidated antioxidant mechanisms by which genistein retards cellular aging. Genistein treatment of UVB-irradiated HDFs in vitro was shown to inhibit elevated intracellular MDA and to increase the activity of intracellular SOD. Furthermore, genistein treatment significantly decreases the number of copies of large deletion mutations in mitochondrial DNA (4977bp and 3895bp, both biomarkers of mitochondrial photoaging in the skin).
Of direct importance in protection from UV-induced skin damage, genistein has been shown to inhibit in vitro chemical and UV-induced DNA oxidation as well as psoralen plus UVA (PUVA) DNA damage. The fact that genistein also reduces erythema and histologic inflammation caused by PUVA may have implications for PUVA therapy by reducing possible short- and long-term adverse reactions.
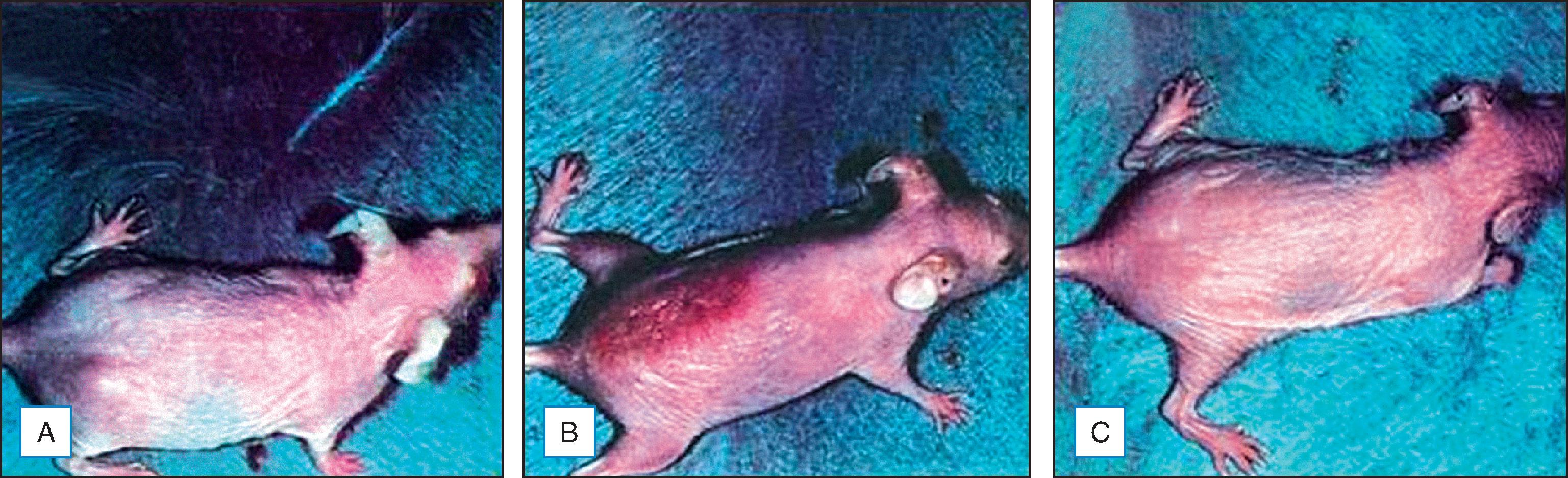
Topical genistein (10 µmol/cm 2 ) protects against acute and chronic UV damage to the skin. After exposure of Skh:1 hairless mice to UVB, topical genistein blocked acute skin burns and inhibited UVB-induced cutaneous wrinkling, as demonstrated clinically in Figs. 25.4 and 25.5 . Histologic analysis confirmed that topical genistein reduces the signs of chronic photodamage—epidermal hyperplasia and reactive acanthosis with nuclear atypia ( Fig. 25.6 ). At a molecular level, UV-induced damage to DNA (as measured by the biomarker 8-hydroxy-2′-deoxyguanosine) was reduced. Also, in Skh:1 mice, topical genistein inhibits UVB-induced pyrimidine dimer formation as well as UVB-generated suppression of repair of solar damage by proliferating cell nuclear antigen (PCNA). This photoprotection by genistein was further demonstrated in vitro with suppression of UVA-induced MMPs responsible for dermal destruction in photoaging. Inhibition of acute UV-induced erythema with topical genistein (5 µmol/cm 2 ) was also demonstrated in humans: topical genistein (applied 30 minutes before UVB) inhibited by one minimal erythema dose (MED) the UVB-induced erythema as shown in Fig. 25.7 . In other recent studies this photoprotection by topical genistein applied pre-UV exposure to rats was confirmed by observation of reduced UVB-induced wrinkling. Pretreatment of human keratinocytes in vitro suppressed the expression of proinflammatory cytokines and proteins released by UVB-treated keratinocytes. Thus topical genistein may protect human skin against photodamage.