Melanocytic nevi represent benign clonal proliferations of the melanocytes in the skin that usually remain stable in size and behavior or disappear during life. Infrequently, melanocytic nevi undergo malignant transformation to melanoma. Understanding molecular and cellular mechanisms underlying oncogene-induced senescence should help identify pathways underlying melanoma development, leading to the development of new strategies for melanoma prevention and early detection.
Key points
- •
Melanocytic nevi share oncogenic molecular mutations with melanomas.
- •
Oncogene-induced senescence explains in part why most nevi are stable and do not undergo progression to malignant melanoma.
- •
Determining mechanisms that reverse melanoma cells to a senescent phenotype could lead to the identification of therapeutic adjuncts in advanced melanoma.
| ANRIL | Antisense noncoding RNA that is involved in chromatin remodeling, transcription, and post-transcriptional processing. It is often abnormally expressed in cancer. |
| Atg7 | Essential autophagy gene, autophagy-related-7, promotes melanoma by limiting oxidative stress and overcoming senescence; inhibition may be of therapeutic value. |
| Bmi-1 | Polycomb complex protein, member of PRC1, which regulates cell cycle inhibitor genes (p16). Its overexpression may promote tumor invasion and metastasis. |
| BMP (bone morphogenic protein) | Member of the transforming growth factor-beta (TGF-beta) superfamily that are involved in proliferation, apoptosis, differentiation, chemotaxis, and angiogenesis. It has been shown to have potent antitumor activity in the skin. In melanoma, it is overexpressed and is thought to promote cell invasion and migration. |
| BMP-Smad1 | This signal and its regulation by epigenetic alterations are significant in Ras-induced senescence |
| BRAF | A member of the RAF family of serine/threonine-specific kinases that is frequently mutated in human melanoma and is a molecular target for therapy |
| BRAF V600E | The V600E mutation results in an amino acid substitution of valine (V) to glutamic acid (E) at position 600 of BRAF. |
| EED (embryonic ectoderm development) | Component of PRC2 |
| EZH2 (enhancer of zeste homolog 2) | Epigenetic modifier and catalytic component of the polycomb repressive complex 2 (PRC2), which is thought to promote growth and metastasis of melanoma. Increased expression is associated with uncontrolled proliferation in melanoma. |
| HRAS G12V | Oncogenic Ras protein that is frequently mutated in cancers. When the amino acid glycine G replaced with amino acid valine V at codon 12, it becomes permanently activated within the cell (proto-oncogene), leading to uncontrolled cell division and tumor formation. |
| JMJD3 | Histone demethylase that promotes melanoma progression and metastasis through regulation of NF-kappa B and BMP signaling |
| MAPK (MAP kinase signaling pathway) | Responsible for relaying extracellular signals from cell membrane to nucleus. Dysregulation of this pathway due to activating mutations in BRAF, RAS and other genes leads to increased signaling activity leading to cell proliferation, invasion, metastasis, migration, survival, and angiogenesis. |
| NRAS | Member of the RAS family of GTPases (small GTPase proteins) that mediate growth factor receptor signaling and are critical for cell proliferation, survival, and differentiation. Activating mutations in NRAS proto-oncogene, particularly at codon 61, are also common in human melanoma |
| OIS | Oncogene-induced senescence |
| p16 (also p16 INK4A ) | Tumor suppressor protein that functions as a cyclin-dependent kinase inhibitor and is encoded by the CDKN2A gene. Plays a significant role in cell cycle regulation; tumor suppressor implicated in the prevention of melanoma and many other cancers. It is 1 of the genes associated with hereditary melanoma and plays a role in cell senescence. |
| p21 WAF1 | Cyclin-dependent kinase inhibitor that mediates p53-dependent cell cycle arrest and likely plays a role as a tumor suppressor. This protein also inhibits apoptosis and may promote cell proliferation in some tumors. |
| p53 | Mutation of this tumor suppressor gene is common in melanoma, more so in many other cancers. In normal cells, p53 plays a role in cell cycle arrest and DNA repair or apoptosis, and can mediate cellular senescence. |
| PcG | Polycomb group |
| PTEN (Phosphatase and tensin homolog) | Tumor suppressor gene that is frequently lost/inactivated in melanoma. The PTEN protein is a phosphatase that negatively regulates the PI3K/Akt pathway and influences cell adhesion, migration, and invasion. |
| PI3K-AKT pathway | Activation of this pathway is one of the most significant signaling pathways in melanoma. It plays a role in melanoma initiation and resistance to therapeutics. |
| PRC | Polycomb repressive complexes 1 and 2 (PRC1 and PRC2), which are protein complexes associated with chromatin condensation and transcriptional repression (epigenetic modifications) PRC1 catalyzes the ubiquitylation of histone H2A and PRC2 methylates H3K27. |
| RB | Tumor suppressor gene that normally arrests cells in the G1 or G1/S phase of the cell cycle by acting as a transcriptional repressor. Loss or inactivation can lead to uncontrolled cell proliferation |
| SAHF | Senescence-associated heterochromatic foci-Domains of condensed chromatin, or heterochromatin that form often in senescent human cells. They play a role in repressing proliferation-promoting genes and their detection can help identify senescent cells. |
| SA β-gal | Senescence-associated β-galactosidase activity that is detectable at pH 6.0 in cells undergoing replicative or induced senescence that us absent in proliferating cells. This is the most commonly used biomarker for senescence. |
| SETDB1 | Histone methyltransferase that is overexpressed in melanoma and accelerates its onset in zebrafish melanoma models harboring the BRAF V600E mutation |
| SMAD1 | Gene that encodes a protein involved in the downstream signaling pathway of BMP |
| SUZ12 | Component of the PRC2 complex |
Melanocytic nevi
A melanocytic nevus is a benign clonal proliferation of melanocytes, the pigment-producing cells of the epidermis, hair follicle, and uveal tract of the eye. Melanocytes are normally interspersed as single cells among keratinocytes in human skin, resting atop the basement membrane. In melanocytic nevi, they are present in greater concentrations, either singly or in adherent nests or clusters of 3 of more melanocytic cells. The clinical appearances of melanocytic nevi are heterogeneous, associated in part with when during life the nevus is acquired and probably also due to the specific differentiation state of the cell of origin and their acquired genetic mutations (see later discussion). They can be considered in 2 major groups: congenital and acquired melanocytic nevi.
A congenital nevus is present in 1% to 3% of neonates at birth and shortly thereafter. They are categorized according to their size (small <1.5 cm, medium 1.5–20 cm, large >20 cm– 40 cm, giant>40 cm). Congenital nevi tend to have a globular pattern on dermoscopy and terminal hair follicles. Histologically, congenital nevi consist of big melanocytes that are fusiform, epithelioid, balloon, or neuroid in shape, tracking down from large nests between collagen bundles along cutaneous appendages, vessels, and nerves. They extend deep into the reticular dermis and the subcutis. It is postulated that these nevi are a result of clonal proliferation of a melanoblast during embryogenesis.
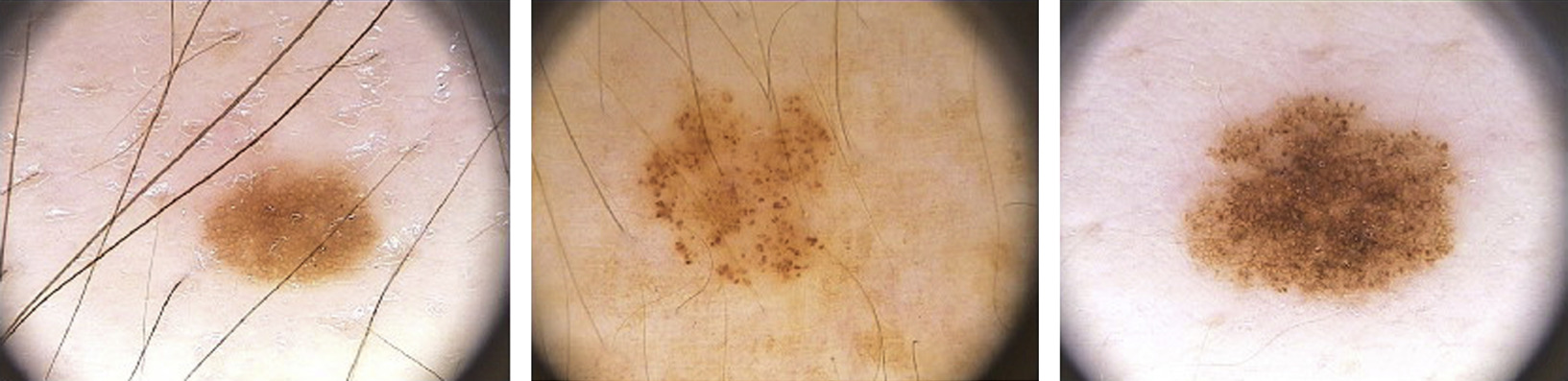
Acquired nevi appear early in childhood, after the first year of life, and increase in number with age, peaking during the third or fourth decade of life. Heredity and environment (ultraviolent radiation) are predisposing factors. Clinically, they are flat, superficial, and horizontally oriented lesions that are usually smaller (<6 mm in diameter) than congenital nevi. Dermatoscopically, they can have a reticular, globular, or homogeneous appearance, alone or in combination ( Fig. 1 ). The distinct patterns probably correspond to different arrangements of the nested an adherent lesional melanocytes. Histologically, melanocytes in acquired nevi usually do not involve the reticular dermis or the subcutis, and the melanocytes are monomorphous and small with an oval shape.
Most melanocytic nevi either disappear or remain stable during life, with fewer than 5% undergoing detectable change when closely monitored. However, it is estimated that 25% to 50% of cutaneous melanomas arise from melanocytic nevi as precursor lesions. A recent meta-analysis by Lin and colleagues of 13 studies involving more than 4000 cases revealed that 32% of melanomas are associated with a nevus. Risk of malignant transformation is associated with increased size of congenital nevi, and risk of cutaneous melanoma correlates with number of total and clinically dysplastic nevi.
In contrast to their malignant counterparts, most melanocytic nevi are benign tumors that initially proliferate but remain stable or disappear during life. However, despite these differences in behavior, nevi and melanomas share somatic mutations in common. A high proportion of large and giant congenital nevi have activating mutations at codon 61 of NRAS , 1 of the 3 major isoforms of the RAS family of GTPase proteins involved in cell growth, survival, and differentiation. Small and medium-sized congenital melanocytic nevi, as well as many acquired melanocytic nevi, contain a key mutation in the BRAF gene, resulting in substitution of glutamic acid for valine at position V600 of the protein within the kinase domain in exon 15. This mutation results in constitutively active BRAF V600E . BRAF is a serine-threonine kinase that is activated by the RAS family of proteins that, when activated, triggers the MAP kinase signaling pathway (MAPK) signaling cascade. Up to 80% of benign nevi carry the BRAF V600E mutation. Both mutations are found at high frequency in cutaneous melanomas, with the BRAF V600E mutation being detected in about 60% to 70% of malignant melanomas. Initial correlation of somatic mutations in nevi with dermatoscopic pattern suggests that BRAF mutations may be most closely associated with globular, rather than reticular, melanocytic nevi. Despite activation of the MAPK pathway, which mediates a potent proliferative signal, benign nevi lose all proliferative activity. The paradox of stable melanocytic proliferations exhibiting oncogenic mutations at high frequency led to the suggestion that melanocytic nevi represent the outcome of oncogene-induced senescence (OIS) in the skin.
Melanocytic nevi
A melanocytic nevus is a benign clonal proliferation of melanocytes, the pigment-producing cells of the epidermis, hair follicle, and uveal tract of the eye. Melanocytes are normally interspersed as single cells among keratinocytes in human skin, resting atop the basement membrane. In melanocytic nevi, they are present in greater concentrations, either singly or in adherent nests or clusters of 3 of more melanocytic cells. The clinical appearances of melanocytic nevi are heterogeneous, associated in part with when during life the nevus is acquired and probably also due to the specific differentiation state of the cell of origin and their acquired genetic mutations (see later discussion). They can be considered in 2 major groups: congenital and acquired melanocytic nevi.
A congenital nevus is present in 1% to 3% of neonates at birth and shortly thereafter. They are categorized according to their size (small <1.5 cm, medium 1.5–20 cm, large >20 cm– 40 cm, giant>40 cm). Congenital nevi tend to have a globular pattern on dermoscopy and terminal hair follicles. Histologically, congenital nevi consist of big melanocytes that are fusiform, epithelioid, balloon, or neuroid in shape, tracking down from large nests between collagen bundles along cutaneous appendages, vessels, and nerves. They extend deep into the reticular dermis and the subcutis. It is postulated that these nevi are a result of clonal proliferation of a melanoblast during embryogenesis.
Acquired nevi appear early in childhood, after the first year of life, and increase in number with age, peaking during the third or fourth decade of life. Heredity and environment (ultraviolent radiation) are predisposing factors. Clinically, they are flat, superficial, and horizontally oriented lesions that are usually smaller (<6 mm in diameter) than congenital nevi. Dermatoscopically, they can have a reticular, globular, or homogeneous appearance, alone or in combination ( Fig. 1 ). The distinct patterns probably correspond to different arrangements of the nested an adherent lesional melanocytes. Histologically, melanocytes in acquired nevi usually do not involve the reticular dermis or the subcutis, and the melanocytes are monomorphous and small with an oval shape.
Most melanocytic nevi either disappear or remain stable during life, with fewer than 5% undergoing detectable change when closely monitored. However, it is estimated that 25% to 50% of cutaneous melanomas arise from melanocytic nevi as precursor lesions. A recent meta-analysis by Lin and colleagues of 13 studies involving more than 4000 cases revealed that 32% of melanomas are associated with a nevus. Risk of malignant transformation is associated with increased size of congenital nevi, and risk of cutaneous melanoma correlates with number of total and clinically dysplastic nevi.
In contrast to their malignant counterparts, most melanocytic nevi are benign tumors that initially proliferate but remain stable or disappear during life. However, despite these differences in behavior, nevi and melanomas share somatic mutations in common. A high proportion of large and giant congenital nevi have activating mutations at codon 61 of NRAS , 1 of the 3 major isoforms of the RAS family of GTPase proteins involved in cell growth, survival, and differentiation. Small and medium-sized congenital melanocytic nevi, as well as many acquired melanocytic nevi, contain a key mutation in the BRAF gene, resulting in substitution of glutamic acid for valine at position V600 of the protein within the kinase domain in exon 15. This mutation results in constitutively active BRAF V600E . BRAF is a serine-threonine kinase that is activated by the RAS family of proteins that, when activated, triggers the MAP kinase signaling pathway (MAPK) signaling cascade. Up to 80% of benign nevi carry the BRAF V600E mutation. Both mutations are found at high frequency in cutaneous melanomas, with the BRAF V600E mutation being detected in about 60% to 70% of malignant melanomas. Initial correlation of somatic mutations in nevi with dermatoscopic pattern suggests that BRAF mutations may be most closely associated with globular, rather than reticular, melanocytic nevi. Despite activation of the MAPK pathway, which mediates a potent proliferative signal, benign nevi lose all proliferative activity. The paradox of stable melanocytic proliferations exhibiting oncogenic mutations at high frequency led to the suggestion that melanocytic nevi represent the outcome of oncogene-induced senescence (OIS) in the skin.
Related posts:
 Basic Science Insights into Clinical Puzzles
Basic Science Insights into Clinical Puzzles
 Establishing Tolerance to Commensal Skin Bacteria
Establishing Tolerance to Commensal Skin Bacteria
 Dermatologic Manifestations of Monogenic Autoinflammatory Diseases
Understanding Inherited Cylindromas
Dermatologic Manifestations of Monogenic Autoinflammatory Diseases
Understanding Inherited Cylindromas
 Mosaic Disorders of the PI3K/PTEN/AKT/TSC/mTORC1 Signaling Pathway
Interleukin-22 and Cyclosporine in Aggressive Cutaneous Squamous Cell Carcinoma
Mosaic Disorders of the PI3K/PTEN/AKT/TSC/mTORC1 Signaling Pathway
Interleukin-22 and Cyclosporine in Aggressive Cutaneous Squamous Cell Carcinoma
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


