Fig. 7.1
Major Th2-related cytokines. Th2 cells secrete various cytokines. IL-4 and IL-13 promote Th2 differentiation, IgE generation, and inflammation and disrupt epidermal barrier function. IL-5 is implicated in eosinophil regulation. IL-31 is associated with pruritus and inflammation
Despite these findings, chronic skin lesions have been shown to be mediated by immunity biased toward Th1 [10]. Th17 cells, which play an essential role in the pathogenesis of psoriasis, may also contribute to the acute phase of AD [11, 12], although this does not appear as prominent as in psoriasis [13]. The role of Th17 immunity in the pathogenesis of AD is uncertain, but IL-17 is hypothesized to enhance the development of AD and prolong exaggeration of skin symptoms [11].
7.2.2 TSLP
TSLP is a member of the IL-7-like cytokines and is expressed by epidermal keratinocytes, mast cells, dendritic cells (DCs), and fibroblasts [14]. TSLP is now receiving attention as an important player in the pathogenesis of AD.
Elevated levels of TSLP are observed in both acute and chronic skin lesions of AD but not in contact allergic dermatitis from nickel or lupus erythematosus lesions [14]. In addition, levels of TSLP expression in the stratum corneum correlate with the Severity Scoring of AD (SCORAD) index [15].
Allergen injection into the skin in individuals with AD causes rapid dermal TSLP expression [16], with Th2 cytokines (IL-4 and IL-13) in conjunction with pro-inflammatory cytokines (tumor necrosis factor [TNF]-α or IL-1α) inducing TSLP production from human keratinocytes [17]. TSLP generation in keratinocytes is also induced by innate immune responses mediated by Toll-like receptor 2/6 and 5 signals [18–20] and protease-activated receptor (PAR)-2 activation through the proteolytic activity of house dust mite allergen [21]. In addition, periostin released from fibroblasts stimulated by IL-4 and IL-13 induces the expression of keratinocyte TSLP [22]. Even more importantly, epidermal TSLP expression is simply stimulated by skin injury or barrier disruption [23, 24].
A crucial role of TSLP in the pathogenesis of AD is in the promotion of Th2 immune responses. Activation of naive CD4 (+) T cells with TSLP induces IL-4 gene transcription, which in turn upregulates TSLP receptor (TSLPR) expression on those cells. This positive feedback mechanism further promotes Th2 cell differentiation [25–27].
TSLP induces the activation and migration of CD11c (+) DCs. These DCs, when primed with specific antigen in the absence of IL-12, promote Th2 cell differentiation from naive CD4 (+) T cells [14, 28]. In addition, TSLP stimulates DCs to produce Th2 cell-attracting chemokines, such as CCL17/thymus and activation-regulated chemokine (TARC) and CCL22/macrophage-derived chemokine (MDC) [14]. Production of CCL17/TARC and CCL22/MDC is increased in AD patients compared with healthy donors. In addition, serum levels correlate with disease activity [29–31]. CCL17/TARC appears to be generated not only by TSLP-primed DCs but also by activated blood mononuclear cells, endothelial cells, and epidermal keratinocytes [32–34]. CCL22/MDC is secreted by macrophages and DCs [35].
A recent study indicated that pruritus is directly provoked by TSLP through TSLPR on transient receptor potential cation channel subfamily A member 1 (TRPA-1)-positive peripheral sensory nerve fibers [36]. Scratching in turn induces epidermal keratinocytes to express TSLP, forming the vicious cycle of AD [24].
7.2.3 IL-31
Activated Th2 cells produce IL-31 [6]. IL-31 is highly expressed in the lesional skin of AD, and serum levels of IL-31 correlate with the clinical severity of AD [37, 38].
IL-31 is a pruritogenic cytokine [39] but also induces skin inflammation. Cutaneous injection of IL-31 into the skin of mice evokes inflammatory cell infiltration. Cell accumulation induced by IL-31 may be partly explained by the fact that epidermal keratinocytes produce CCL17/TARC and CCL22/MDC in response to IL-31 [39].
7.2.4 IL-18 and IL-33
IL-18 and IL-33 belong to the IL-1 family of cytokines. IL-18 has been generally known as a Th1 cytokine but also contributes to Th2 immunity. IL-18 stimulates basophils, mast cells, and CD4 (+) T cells to produce IL-4 and IL-13 [40–42]. AD patients display elevated serum concentrations of IL-18, with these levels correlating with disease severity and serum IgE [43].
7.3 Mast Cells
Mast cells are tissue-resident inflammatory cells that possess metachromatic cytoplasmic granules. Mast cells express a variety of cell surface receptors including stem cell factor (SCF) receptor CD117 (also known as c-kit) and a high-affinity IgE receptor, Fc epsilon receptor I (FcεRI) [49, 50].
Increased numbers of dermal mast cells are observed in skin lesions of AD [51]. Mast cells show various stages of degranulation in acute skin lesions but are fully granulated in chronic skin lesions [3]. However, whether and how mast cells contribute to the pathogenesis of AD remains unclear [52].
Th2 skewing in AD may affect mast cell proliferation and activation. IL-4 enhances the IgE-induced upregulation of FcεRI on mast cells [53]. IL-5 promotes the proliferation of mast cells in conjunction with SCF [54]. In contrast, the Th1-related cytokine IFN-γ decreases mast cell numbers [50].
Mast cells activated by IgE plus antigens produce and secrete three categories of substances: (1) chemical and protein mediators (histamine, serotonin, heparin, proteases, and even major basic protein [MBP]) [55]; (2) a wide variety of cytokines (e.g., IL-3, IL-4, IL-5, IL-6, and IL-13), chemokines (e.g., CCL3/macrophage inflammatory protein [MIP]-1α, CCL4/MIP-1β, CCL5/regulated on activation, normal T cell expressed and secreted [RANTES], and CXCL10/IFNγ-induced protein-10 [IP-10]), and growth factors (e.g., SCF); and (3) lipid mediators (prostaglandins [PGs], leukotrienes [LTs], etc.).
Histamine is a representative pruritogen released by mast cells. In addition, mast cells produce a tryptase that cleaves and activates its receptor PAR-2 on nerve fibers, provoking pruritus [56].
Mast cell-derived histamine appears to play important roles in tissue remodeling [51]. Synthesis of collagen and periostin by fibroblasts is induced by histamine. Periostin, in turn, stimulates fibroblast collagen synthesis [57] and also induces keratinocyte proliferation [58].
Mast cells are capable of generating Th2-related cytokines (IL-4, IL-5, IL-13, and IL-13), thereby contributing to Th2 immunity. In skin lesions of AD, a high proportion of dermal mast cells express IL-31 [59].
PGD2 is one of the cyclooxygenase metabolites of arachidonic acid. Mast cells are one of major producers of PGD2 in the skin. Chemoattractant receptor-homologous molecule on Th2 cells (CRTH2), a receptor for PGD2, is expressed on Th2 cells, eosinophils, basophils, and type 2 innate lymphoid cells (ILC2) [60, 61]. CRTH2 provides stimulatory signals in those cells and exerts chemotactic activity. CRTH2 has been shown to play an essential role in mouse models of AD [62].
7.4 Eosinophils
Eosinophils are granulocytes containing two major types of granules: specific and primary. Specific granules contain cationic proteins such as eosinophil MBP, eosinophil peroxidase (EPO), eosinophil cationic protein (ECP), and eosinophil-derived neurotoxin (EDN). These cationic proteins are protective against bacteria, parasites, and/or viruses but are also toxic to tissue cells [63]. Primary granules show enriched levels of Charcot-Leyden crystal protein (galectin-10) [7, 50]. Eosinophils exert their function by releasing pro-inflammatory mediators, including these proteins, cytokines (e.g., TNF, transforming growth factor [TGF], IL-1, IL-3, IL-4, IL-5, IL-8, IL-10, and IL-13), and eicosanoids (e.g., LTC4, LTD4, and LTE4) [64, 65].
In histology, eosinophils are usually seen in the acute phase of AD but are barely observed in chronic lichenified lesions. However, significant deposition of eosinophil MBP can be detected in the dermis. Tissue eosinophilia is often associated with blood eosinophilia and correlates with the severity of AD [66].
The Th2-related cytokine IL-5 is a key cytokine for eosinophils, and elevated levels of IL-5 mRNA are detected in the skin lesions of AD [67]. IL-5 is produced mainly by stromal cells in bone marrow and by Th2 cells. In addition to IL-5, eotaxins (CCL11/eotaxin and CCL26/eotaxin-3), IL-3, and granulocyte-macrophage colony-stimulating factor (GM-CSF) also promote the development and differentiation of eosinophils from CD34 (+) hematopoietic progenitor cells [68]. Eosinophils that have developed in the bone marrow then mobilize into the blood circulation in response to IL-5 and/or eotaxin stimulation [69, 70] and migrate to inflamed tissues where Th2 immune mechanisms predominate. IL-4 and IL-13 induce P-selectin and endothelial cell vascular cell adhesion molecule 1 (VCAM-1) expression on dermal endothelial cells [71, 72]. Endothelial P-selectin promotes eosinophil rolling on endothelial cells via P-selectin glycoprotein ligand-1 (PSGL-1) [73]. In addition, since very late antigen-4 (VLA-4), a ligand for VCAM-1, is expressed on eosinophils, but not on neutrophils, the IL-4/IL-13-VLA-4 axis may represent a key step in eosinophilic inflammation [74]. IL-4 and IL-13 stimulate dermal fibroblasts to produce eotaxins [75, 76]. Eotaxins and the Th2-related protein TSLP exert chemoattractant activity for eosinophils [77].
On the other hand, IFN-γ, a Th1 cytokine, stimulates dermal fibroblasts to produce CCL5/RANTES [78] and ecalectin (galectin-9) [79], which are both eosinophil chemoattractants [80, 81]. Eosinophils express CXCR3 and exhibit chemotaxis to CXCL10/IP-10 and CXCL9/monokine induced by IFN-γ (Mig) [82]. Importantly, eosinophils themselves generate a Th1 cytokine, IL-12 [83]. Eosinophils may thus participate in the immune reactions of not only Th2-type inflammation but also Th1-type inflammation, such as chronic lesions of AD (Fig. 7.2).
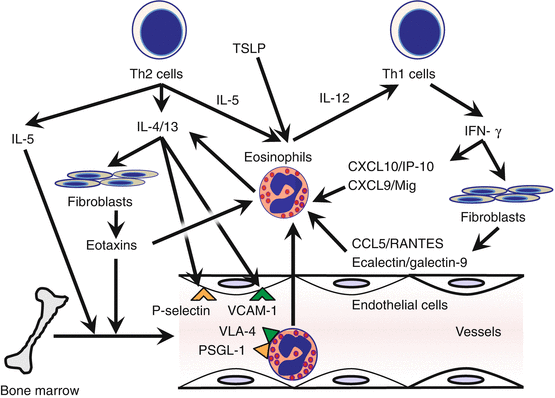
Fig. 7.2
Eosinophil migration into inflamed skin. IL-5 mobilizes the eosinophil pool from bone marrow into the peripheral circulation. The Th2 cytokines IL-4/IL-13 stimulate fibroblasts to generate eosinophil chemokine eotaxins and endothelial cells to express the cell adhesion molecules P-selectin and VCAM-1. The Th1 cytokine IFN-γ mediates CXCL10/IP-10, CXCL9/Mig, CCL5/RANTES, and ecalectin/galectin-9 production, leading to eosinophil accumulation in inflammatory lesions biased toward Th1. IP-10, IFNγ-induced protein-10; Mig, monokine induced by IFN-γ; PSGL-1, P-selectin glycoprotein ligand-1; RANTES, regulated on activation, normal T cell expressed and secreted; VCAM-1, vascular cell adhesion molecule 1; VLA-4, very late antigen-4
The pathogenic roles of eosinophils in skin inflammation have been a focus of debate, and whether toxic granule proteins (ECP, EDN, EPO, and MBP) derived from eosinophils cause tissue damage remains contentious. One study has demonstrated that dermal eosinophilia induced by intradermal CCL5/RANTES injection did not provoke macroscopic changes in the skin of atopic patients [84]. More importantly, the monoclonal IL-5 antibody mepolizumab has been found to ameliorate only peripheral eosinophilia, not other AD symptoms [85].
A number of recent lines of evidence have suggested a possible contribution of eosinophils to tissue remodeling. TGF-β1 derived from eosinophils promotes fibroblast proliferation, collagen synthesis, and lattice contraction [86]. Eosinophils also produce metalloproteinase 9, vascular endothelial growth factor, basic fibroblast growth factor, and angiogenin [87–89]. In AD patients, intradermal allergen challenge has been found to induce infiltration of TGF-β1 (+) eosinophils, α-smooth muscle action (+) myofibroblasts, and procollagen-I (+) cells [90], which was inhibited by anti-IL-5 monoclonal antibody and TGF-β1-neutralizing antibody [90, 91].
In a murine model of prurigo reactions, the presence of dermal eosinophils correlated with scratching behavior [92]. In a model of contact toxicant reactions, skin innervation and scratching behaviors were dependent on eosinophils [93]. Eosinophils promote branching of sensory neurons in vitro [94]. In humans, MBP and EPO stimulate mast cells to release histamine through Mas-related gene (Mrgx2) [95]. Eosinophils may thus participate in the development of intractable itch in AD.
7.5 Basophils
Basophils constitute less than 1% of peripheral blood leukocytes and have a half-life of 1–3 days [50]. Basophils share some functional and morphological similarities with mast cells (e.g., expression of the high-affinity IgE receptor FcεRI and secretion of histamine). These similarities initially led to the idea that basophils were minor and redundant relatives or blood-circulating precursors of tissue-resident mast cells [96]. However, recent accumulated evidence has indicated nonredundant roles of basophils compared to mast cells in innate and acquired immunity. In a number of skin diseases, basophils have been demonstrated to be present in skin lesions [97].
With regard to the hematopoiesis of human basophils, a close lineage relationship with eosinophils has recently been demonstrated, as seen in the presence of immature basophils with a basophil-eosinophil hybrid phenotype and the capacity to generate eosinophil-specific MBP [98].
Both TSLP and IL-3 promote the development and maturation of basophils from bone marrow progenitor cells [99]. The Th2 cytokine IL-4 induces expression of VCAM-1. Like eosinophils, human basophils express the VCAM-1 ligand VLA-4 [74]. In addition, P- and E-selectin also contribute, at least partially, to the adhesion of human basophils to endothelium [100]. In mice, functional PSGL-1, modified by α(1,3) fucosyltransferases (FTs)-IV and (FT)-VII, and L-selectin play roles in the initial recruitment of basophils in chronic allergic inflammation [101]. CCL11/eotaxin, CCL2/monocyte chemotactic protein (MCP)-1, CCL13/MCP-4, and histamine (through the histamine H4 receptor) are thought to be chemoattractive for basophils [102–104].
IL-3 from activated T cells promotes basophil development [105]. When activated by FcεRI cross-linking, IL-3-elicited basophils degranulate and release histamine, leukotrienes (e.g., LTB4, LTD4, LTE4), PGs (e.g., PGD2 and PGE2), and cytokines (e.g., IL-4 and IL-13) [106–108]. TSLP, a key player in Th2 immunity of AD, also regulates basophil development and peripheral basophilia [99]. TSLP-elicited basophils respond to IL-3, IL-18, and IL-33 and produce IL-4 [99]. Of note, the amounts of secreted cytokines are larger than from IL-3-elicited basophils.
Basophils are considered to function as antigen-presenting cells (APCs) in mice [109]. However, human basophils have not yet been confirmed to possess APC activity [110–112].
Basophil infiltration is observed in the lesional skin of AD patients [97]. Although a few basophils are present in AD lesions, patch testing with house dust mite antigen has been found to result in significant basophil infiltration into the dermis and even epidermis within 48 h after exposure [97]. In allergen-induced late-phase reactions of AD, maximal eosinophil infiltration occurred at 6 h, whereas peak basophil accumulation was seen at 24 h, one-third of which had morphologic appearances suggestive of degranulation [113, 114].
In a murine model of IgE-dependent eosinophilic dermatitis, infiltration of eosinophils was seen to be dependent on basophils [115]. In IgE-mediated chronic allergic inflammation (IgE-CAI) [116], basophils are indispensable for the induction of eosinophils and other inflammatory cells, even though dermal basophils constituted only 2% of total infiltrative cells [92, 116]. Based on these findings, we can assume that basophils play important roles in the pathogenesis of human AD, despite constituting only a minority population among inflammatory cells. However, whether and how basophils play a pathogenetic role in AD remains to be determined and warrants a focus on elucidation in the near future.
Conclusions
Th2-type immunity represents a key inflammatory process in the pathogenesis of AD. Th1 and Th17 immunity also appear to be involved in inflammation. Mast cells and eosinophils participate in not only inflammatory processes but also in pruritus and tissue remodeling. Further research is needed to clarify the actual pathogenetic roles of basophils in AD.
References
2.
Kabashima K. New concept of the pathogenesis of atopic dermatitis: interplay among the barrier, allergy, and pruritus as a trinity. J Dermatol Sci. 2013;70(1):3–11.PubMed
4.
Palmer CN, Irvine AD, Terron-Kwiatkowski A, Zhao Y, Liao H, Lee SP, et al. Common loss-of-function variants of the epidermal barrier protein filaggrin are a major predisposing factor for atopic dermatitis. Nat Genet. 2006;38(4):441–6.PubMed
5.
Howell MD, Kim BE, Gao P, Grant AV, Boguniewicz M, Debenedetto A, et al. Cytokine modulation of atopic dermatitis filaggrin skin expression. J Allergy Clin Immunol. 2007;120(1):150–5.PubMedPubMedCentral
6.
Sonkoly E, Muller A, Lauerma AI, Pivarcsi A, Soto H, Kemeny L, et al. IL-31: a new link between T cells and pruritus in atopic skin inflammation. J Allergy Clin Immunol. 2006;117(2):411–7.PubMed
7.
Simon D, Braathen LR, Simon HU. Eosinophils and atopic dermatitis. Allergy. 2004;59(6):561–70.PubMed
8.
Werfel T, Allam JP, Biedermann T, Eyerich K, Gilles S, Guttman-Yassky E, et al. Cellular and molecular immunologic mechanisms in patients with atopic dermatitis. J Allergy Clin Immunol. 2016;138(2):336–49.PubMed
9.
Simpson EL, Bieber T, Guttman-Yassky E, Beck LA, Blauvelt A, Cork MJ, et al. Two phase 3 trials of dupilumab versus placebo in atopic dermatitis. N Engl J Med. 2016;375(24):2335–48.PubMed
10.
Grewe M, Gyufko K, Schöpf E, Krutmann J. Lesional expression of interferon-gamma in atopic eczema. Lancet. 1994;343(8888):25–6.PubMed
11.
Koga C, Kabashima K, Shiraishi N, Kobayashi M, Tokura Y. Possible pathogenic role of Th17 cells for atopic dermatitis. J Invest Dermatol. 2008;128(11):2625–30.PubMed
12.
Noda S, Suárez-Fariñas M, Ungar B, Kim SJ, de Guzman Strong C, Xu H, et al. The Asian atopic dermatitis phenotype combines features of atopic dermatitis and psoriasis with increased TH17 polarization. J Allergy Clin Immunol. 2015;136(5):1254–64.PubMed
13.
Guttman-Yassky E, Lowes MA, Fuentes-Duculan J, Zaba LC, Cardinale I, Nograles KE, et al. Low expression of the IL-23/Th17 pathway in atopic dermatitis compared to psoriasis. J Immunol. 2008;181(10):7420–7.PubMedPubMedCentral
14.
Soumelis V, Reche PA, Kanzler H, Yuan W, Edward G, Homey B, et al. Human epithelial cells trigger dendritic cell mediated allergic inflammation by producing TSLP. Nat Immunol. 2002;3(7):673–80.PubMed
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


