JAML–CAR
DETC express JAML (junction adhesion molecule like protein) at low level in steady state and upregulate its expression upon activation. The binding partner of JAML, the coxsackie and adenovirus receptor (CAR), is expressed by keratinocytes. CAR-mediated JAML clustering recruits PI3K, in a similar mechanism as described for CD28-mediated costimulation [156]. PI3K mediates costimulatory function of JAML through the same binding motif found on B7 family members CD28 and ICOS. This leads to proliferation and production of IFN-γ, IL-2 and TNF-α by DETC. The inhibition of this interaction results in reduced γδ T activation and delayed wound healing in the skin. Based on the expression pattern of JAML and CAR in the intestine, their interaction could also play a role in activation of γδ IEL in this organ.
CD100–plexin B2
DETC express CD100/semaphorin 4D (Sema4D), a member of the semaphorin family of molecules with a known role in axonal guidance. CD100 on DETC engages plexin B2 on keratinocytes, and in vitro ligation of CD100 leads to activation of cofilin and ERK activation and the rounding of DETC. DETC from CD100 deficient mice show delayed rounding, which correlates with a wound healing defect in these mice [157]. In the intestine, all IEL express CD100 and epithelial cells express plexin B2. CD100-deficient mice show exacerbated inflammation and tissue damage in the DSS-colitis model. CD100 deficiency renders γδ IEL unable to produce KGF-1, and KGF-1 administration ameliorates intestinal tissue damage in these mice [93]. Collectively these data indicate the importance of CD100 in mediating tissue homeostasis and repair by DETC.
Aryl hydrocarbon receptor (AhR)
The aryl hydrocarbon receptor (AhR) is a member of the basic helix-loop-helix (HLH)/Per-Arnt-Sim (PAS) superfamily known for their roles in sensing of environmental factors. It is a ligand-activatable transcription factor, and regulates several xenobiotic-metabolizing enzymes. Cell-intrinsic expression of AhR is important for the maintenance of DETC in the skin. AhR-deficient DETC have altered morphology and do not extend dendrites to contact neighbouring cells. Although AhR deficiency does not impair thymic generation or export of these cells to the skin, their numbers in the epidermis decline over time [71]. Recent studies found a similar role for AhR in the maintenance of γδ IEL in the gut [83].
CD27
CD27 is a member of the TNF superfamily and its binding partner CD70 is constitutively expressed on thymic epithelial cells. In the thymus, a CD25+CD27+ DN population contains the precursors that subsequently diverge into the CD27− RORγ+ IL-17 producing lineage and the CD27+ IFN-γ producing lineage. This divergence is stable and largely not interconvertible even in the face of infection in the periphery [114]. CD27−/− mice contain less IFN-γ producing γδ T cells, indicating that CD27 is not only a marker but also a regulator of effector cell fate determination of γδ T cells.
WC1 (Workshop Cluster 1)
WC1 are germline-encoded coreceptors of the scavenger receptor cysteine-rich (SRCR) family of transmembrane glycoprotein, consisting of 11 extracellular SRCR domains. WC1 is related to the CD163 family, with closest homology to CD163c-alpha/CD163L1 or SCART) [56]. WC1 is uniquely expressed by γδ T cells in cattle, camelids and pigs. Gene orthologues have been described in mice and human but no functional gene product has been found. The cytoplasmic domain of WC1 contains ITAMs. Specific tyrosine moieties are constitutively phosphorylated and associate with Src family tyrosine kinases. Phosphorylation of the second tyrosine residue is required for the role of WC1 in potentiating γδ TCR-mediated proliferation in vitro, an observation supporting the role of WC1 as a costimulatory molecules [150].
Bovine γδ T cells are serologically defined based on available antibodies into WC1.1+, WC1.2+ and WC1− subsets. WC1 are not merely markers but define functional subsets of bovine γδ T cells with distinct age-dependent turnover kinetics [151]; [118]; [119]. WC1.1+ proliferate more and produce more IFN-γ in allogeneic MLR in vitro compared to WC1.2+ γδ T cells [118]; [119]. Intranasal BCG vaccination induces higher expansion or recruitment of the WC1.1+ subset into the lung, as well as eliciting higher production of IFN-γ from these cells [110]; [49]. Virus infection induces differential cytokine and chemokine production in the subsets, with WC1.1+ and WC1− cells producing MIP-1α and GM-CSF while WC1.2+ cells producing IL-10 and TGF-β [90].
SCART
SCART, another member of the SCRC superfamily, shares significant homology with WC1 but is not its gene orthologue. The expression of SCART1 and SCART2 are largely identical. In the thymus, SCART2 upregulation is first detected at the DN2 stage and does not require a TCR signal. However, its expression is downregulated by strong TCR ligation. In the adult periphery, SCART2 positivity defines a population of thymic-derived IL-17-producing γδ T cells enriched for Vγ4 reactivity residing in the skin dermis and skin-draining LN [74].
NKG2D
NKG2D interacts with various stress-inducible molecules including MICA, MICB, ULBP, H60, Rae1 and MULT. In γδ T cells, the co-ligation of NKG2D and γδ TCR potentiates γδ T cell activation, supporting a costimulotary role for this molecule. However, NKG2D is also capable of TCR-independent activation of γδ T cells in other conditions.
BTLA (B– and T– Lymphocyte Attenuator)
BTLA, an inhibitory costimulatory molecule of the B7 superfamily, has recently been described to play a role in the homeostasis and regulation of γδ T cells [6]. RORγt represses BTLA while IL-7 signalling increases its surface expression. BTLA expression limits the responsiveness of γδ T cells to IL-7 mediated proliferation thereby regulating its homeostatic number at steady state. BTLA also inhibits the production of IL-17 and TNF from CD27− γδ T cells in vitro. In a γδ T cell-dependent model of dermatitis, BTLA deficiency exacerbates, and BTLA ligation ameliorates disease.
Other costimulators
γδ T cells and αβ T cells share many common costimulatory molecules. However, while costimulators have a qualitative role determining the outcome of αβ T cell response between activation and anergy, the effect of costimulation in γδ T cells appears to be quantitative. For example, CD28 ligation does enhance in vitro response of γδ T cells to TCR crosslinking and to allogeneic DC. However, the expression of CD28 was found to be variable and its importance in the activation of γδ T cells unresolved [54]. In the presence of γδ TCR crosslinking, TNF SF family member CD40L is known to costimulate the proliferation and cytolytic function of thymic γδ T cells [113]. Persistent upregulation of CD30, another TNF SF member, can be induced upon in vitro activation of γδ T cells. CD30 engagement leads to enhanced cytokine and chemokine production [7].
Heterogeneity Among γδ T Cell Subsets
Recent studies strongly suggest that γδ T cells can be divided into an innate or natural population and an adaptive or inducible population [22, 73, 144]. γδ T cell populations that are generated in the foetal thymus typically home to peripheral sites such as the skin and the gut epithelium, where they self-renew in situ. These cells are pre-programmed in the thymus into distinct effector lineages, express markers indicative of partial activation, and at least a subset of them can be activated in a TCR-independent manner. Collectively, these features place them within the innate spectrum of γδ T cells. On the other end of the spectrum are γδ T cells showing more adaptive-like features, typically circulating in the blood, spleen and lymph nodes, where about half the population has a CD44−CD62L+ naive phenotype. There is no evidence of effector pre-programming in these cells. Cytokine production potential is plastic with more delayed kinetics relative to that of their more innate counterparts. These cells have different derivative requirements and can be generated from adult thymic and bone marrow precursors. Recall response can be elicited from these ‘adaptive’ γδ T cells in a number of different species [99, 127, 128].
An Example of Microanatomical Specialisation: Dermal γδ T Cells Versus DETC
Several recent studies have highlighted the microanatomical and functional segregation of two populations of γδ T cells in the mouse skin. Using intravital multiphoton imaging of fluorescently tagged mice (CXCR6-GFP knock-in), two groups have identified a novel population of γδ T cells residing in the dermis [47, 132]. In the epidermis, DETC are sessile and constantly extend and retract their dendritic projections, while dermal γδ T cells show a different morphology and migrate in the dermis at a speed of 2–4 μm/min (Fig. 4.1). A subset of these cells also makes contact with MHC-II+ cells, presumably antigen presenting cells, in the dermis. These behaviours suggest an active role for these cells in immune-surveillance of the skin. While DETC are uniformly Vγ5+, dermal γδ T cells are relatively heterogeneous, expressing Vγ5, Vγ4 or other TCRs [47, 132]. In addition, the two populations also show differential expression of γδ TCR levels, CD25, CD43, CCR6 and NK1.1. Both populations are generated perinatally and homeostatically maintained by self-renewal in situ in an IL-7 dependent manner. In addition, DETC but not dermal γδ T cells are IL-15 dependent [29, 132]. While DETC are thymically pre-programmed by skint1 ligation to produce IFN-γ [114, 138], dermal γδ T cells constitutively express RORγt and IL-23R, and rapidly produce IL-17 following exposure to IL-1β plus IL-23 [16, 47]. In an intradermal model of BCG infection, dermal γδ T cells are the earliest population showing rapid IL-17 production, which mediates recruitment of neutrophils to the site of antigen deposit. In Tcrd −/− mice, the recruitment of neutrophils to the infected ear skin is diminished, and the downstream adaptive response of antigen-specific conventional CD4+ T cells reduced [132]. Nonetheless, these two anatomically segregated of γδ T cells may have functional overlap in certain scenarios, as a recently described subset of DETC is capable of rapidly producing IL-17 in response to wounding and UV irradiation [87, 88].
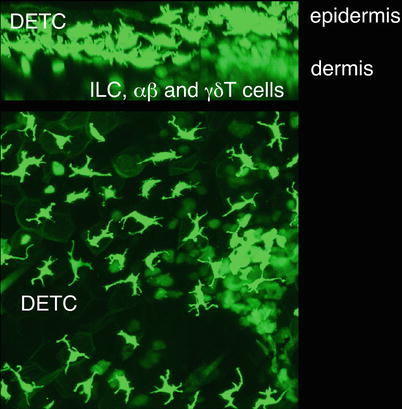
Fig. 4.1
Dendritic epidermal T cells (DETC) and dermal lymphoid cells in the skin of Cxcr6gfp/+ mice. The non-migratory DETC reside in the epidermis, where their cell processes are anchored at squamous keratinocyte tight junctions, giving the cells their characteristic dendritic morphology. Dermal lymphoid cells, which include αβ and γδ T cells as well as innate lymphoid cells, are more amoeboid in morphology and actively migrate throughout the dermis. The image is a z-projection through a volume of 82 μm of ear skin, including epidermis and dermis
Interaction Between γδ T Cells and Other Cell Types
Interaction with Other T Cell Subsets
Mature TCR γδ+ thymocytes are reduced in numbers in Tcrβ-deficient and pTα-deficient, but not Tcrα-deficient mice, indicating a role for the CD4+CD8+ DP αβ T precursors on the development of γδ T cells in the thymus. DP cells are capable of conditioning γδ T precursors via lymphotoxin (LT)-mediated RelA and RelB pathways, regulating the expression of transcription factors RORγt and RORα4 to direct their differentiation into Tγδ17 cells [109, 129]. In Tcrd −/− mice hyperproliferation of the αβ T cell compartment is often seen, suggesting that γδ T cells may compete with αβ T cells for homeostatic cytokines, such as IL-7. During infection, early cytokine production from γδ T cells may directly or indirectly polarise delayed adaptive responses downstream and impact on the efficacy of pathogen clearance. For instance, infection with Listeria monocytogenes or Nippostrongylus brasiliensis elicits rapid production of IFN-γ and IL-4, respectively, from γδ T cells of the peritoneal cavity [35].
In models of autoimmune-mediated inflammation, γδ T cells may exacerbate disease. Tγδ17 that are activated by IL-23 during MOG/CFA-induced experimental autoimmune encephalomyelitis (EAE) inhibit the de novo induction of regulatory T cells (Treg) from conventional T cells and confer resistance of antigen-specific effector T cells towards Treg-mediated suppression [108]. Early cytokine production from resident mucosal γδ T cells aggravates colitis induced by adoptive transfer of effector CD4+ T cells in lymphopenic hosts, which can be inhibited by adoptive transfer of Tregs [159]. Treg and Tγδ17 are also likely to be engaged in dynamic interactions during the steady state, as the reduction in number and function of CD4+CD25+ Treg in Pdk1-deficient mice allows the expansion of pathogenic Tγδ17 cells that drive chronic intestinal inflammation [103].
Interaction with B Cells
Human Vγ9+Vδ2+ cells are the majority population of γδ T cells in the peripheral blood. A subset of these cells has a CCR7+CD62L+ central memory (TCM) phenotype, expresses the B cell follicle-homing chemokine receptor CXCR5 and the costimulatory molecules CD40L and ICOS, and is capable of producing IL-2, IL-4 and IL-10. In vitro, these cells are capable of helper T cell function to drive antibody production and isotype class switching [13]. In patients with genetic lymphopenia, γδ cells remain functional and are capable of providing B cell help in place of CD4+ T cells, leading to the formation of small germinal centres. These patients often show hyperimmunoglobulinemia of IgE, indicative that γδ T cells are capable of driving Ig class switching in vivo [34, 95].
γδ T Cells and Antigen Presenting Cells (APC)
The interactions between γδ T cells and antigen presenting cells (APC) in the thymus and the periphery are crucial for their generation, maintenance and function. In the thymus, Vγ5+ precursors and medullary thymic epithelial cells (mTEC) are mutually dependent. LTi (lymphoid tissue inducer) cells as well as Vγ5+ γδ precursor cells provide RANK ligand signal driving the maturation and expression of Aire in mTEC [116]. While Aire expression per se has no role in the generation of Vγ5+ DETC, mature mTEC express skint1, which is crucial for the programming of effector functions in these cells.
Intravital imaging revealed that γδ T cells at peripheral sites constantly make contacts with resident antigen presenting cells (APCs) [24]. Examples include contact between DETC and Langerhans cells (LC) in the epidermis, and between dermal γδ T cells and MHC-II+ cells in the dermis. While the depletion of LCs does not affect maintenance of DETCs [134], DETCs are producers of XCL1/lymphotactin, a chemokine known to attract DC [8]. While the activation of naïve αβ T cells strictly requires interaction with mature professional APC ie dendritic cells, immature DC have been shown to be able to activate γδ T cells. Vγ9+ T cells engage in reciprocal interactions with DCs, and are capable of potentiating DC maturation via production of cytokines, as well as relieving inhibitory signals that block DC maturation [62, 104, 149].
γδ T cells may themselves play a role as antigen presenting cells (APCs) thereby bridging the early innate immunity to the delayed adaptive T cell-mediated response [12, 97]. Human Vγ9+Vδ2+ cells have been shown to present antigen to CD4+ T cells, and cross-present antigen to CD8+ T cells. Activated Vγ9+ cells display several salient features that enable them to function as professional APC, including uptake of soluble antigens, phagocytosis, upregulation of CCR7 enabling homing to secondary lymphoid organs, upregulation of MHC-II and the costimulatory molecules CD80 and CD86.
Interaction with Other Myeloid Cells
γδ T cells engage with myeloid populations in a bidirectional manner. Efficient presentation of phosphoantigen to Vγ9+ cells in vitro requires the presence of monocytes [94]. Previous studies showed defective maturation of monocytes derived from mice with a deficit in γδ cells [130]. In vivo, different subsets of γδ T cells have protective roles in controlling macrophage-mediated immunopathological tissue damage in Listeria infection. Vγ1+ T cells mediate apoptosis of activated macrophages in a Fas:FasL dependent manner [28]. The interaction between Vγ4+ cells and activated macrophages promotes cytokine and chemokine production by macrophages on one hand, while inducing IL-10 production from Vγ4+ cells themselves on the other [137]. Yet both the Vγ1+ and Vγ4+ subsets are protective against Listeria-induced liver injury in independent adoptive transfers.
Interaction with Innate Lymphoid Cells (ILCs)
Administration of chitin elicits type 2 innate inflammation in the lungs involving ILC2-mediated recruitment of eosinophils and alternatively activated macrophages. When innate lymphoid cells are depleted, there is an enhancement of Tγδ17 activation and prolonged neutrophil influx, suggesting inter-regulation between ILC2 and Tγδ17 in determining the identity of infiltrating myeloid cells in response to this allergen [143].
Trafficking of γδ T Cells
Proper homing and localisation is of paramount importance to the generation and function of γδ T cells. The medullary localisation of Vγ5+ precursors is sensitive to pertussis toxin indicating a requirement for G protein coupled protein in this process. Mice deficient for an enzyme required in synthesising E- and P- selection ligands have reduced number of DETC in the skin despite normal generation of DETC in the thymus indicating the importance of these molecules for their homing and/or maintenance in the skin [69]. In the foetal thymus, TCR-mediated upregulation of CCR10 is required for homing of DETC to the skin [70]. CCR9 deficient mice have a deficiency in gut intraepithelial αβ and γδ T lymphocytes [20], indicating that the same chemokine-receptor is used for targeting both populations of T cells to this site.
Despite tissue-specific localisation of γδ T cells of restricted TCR diversity, the individual TCR per se is not required to dictate their tissue homing specificity. In Vγ4Vδ5 γδ TCR transgenic mice, DETC expressing the transgenic TCR instead of the canonical Vγ5 are generated and home to the epidermis, albeit at a reduced number [10]. However, the expression of a specific TCR in a specific tissue compartment is clearly of physiological relevance. As evidence, Vγ5Vδ1 deficient mice have increased susceptibility to chemically induced carcinogenesis in the skin [131]. The Taconic strain of FVB mouse which harbours heterogeneous rather than the canonical Vγ5+Vδ1+ DETC is more susceptible to spontaneous and induced dermatitis [82].
The activation of γδ T cells initiates modulation of chemokine receptors and acquisition of differential homing potential. Human Vγ9+ cells constitutively express CCR5, which is downregulated as the cells become activated [43]. Activation is associated with the upregulation of CCR6 and CCR7 [11, 14]. In the lungs, intratracheal instillation of LPS or BCG induces an early TLR dependent infiltration of γδ T cells. Infiltration of γδ T cells is dependent on CCL2:CCR2, as recruitment was diminished in CCL2 (MCP-1) knockout mice and after CCL2 neutralisation, and restored with administration of CCL2 [105]. Hepatic Tγδ17 cells have a protective role in chronic liver injury that is independent of IL-17 production but dependent on CCR6-mediated homing to the liver. In two models of chronic liver injury, CCR6−/− mice develop more severe fibrosis, and adoptive transfer of wildtype γδ T cells ameliorates hepatic inflammation and injury [51]. Notably CCR6-mediated homing of Tγδ17 may be associated with very different outcome depending on the target organ. In a cutaneous inflammation model, CCR6 deficiency precludes the development of inflammation and hyperplasia. CCL20:CCR6 interaction is required for the recruitment of γδ T cells to the epidermis, where these cells exacerbate inflammation via production of IL-17 and IL-22 [86].
Memory Responses in γδ T Cells
There are documented instances of rapid mobilisation and expansion of γδ T cells akin to adaptive memory, however clonal expansion is of a relatively lower magnitude compared to a classical memory response of αβ T cells. In addition, although context specificity is evident, demonstration of cognate antigen specificity is lacking, and probably impractical, given the relatively unknown antigen recognition apparatus and less stringent activation requirement of γδ T cells. Nonetheless, protective effect of γδ T cells in secondary infection is a promising venue to be explored for vaccine development.
Oral infection with Listeria monocytogenes elicits the expansion of a population of γδ T cells in mesenteric LN that produce both IL-17 and IFN-γ. These cells are capable of rapid response to secondary oral infection with Listeria but not the unrelated pathogen Salmonella, and to oral but not intravenous infection with L. monocytogenes, demonstrating context specificity in response [128]. Infection of Mycobacterium bovis in cattle induces rapid activation and IFN-γ production from CD8+CD45RO+ γδ T cells that respond more strongly towards M. bovis infected rather than BCG-infected macrophages [57]. Primary infection of macaques with BCG induces polyclonal expansion of Vγ9+Vδ2+ cells in the blood. These cells rapidly expand upon secondary BCG infection, and expansion is associated with clearance of bactereamia in the blood. More importantly, in BCG-immune macaques these cells also expand in response to secondary infection with M. tuberculosis and showed protection against this usually fatal infection [127]. Within the skin, we have observed an increase in the number of dermal γδ T cells with age, suggesting that these cells may also be capable of exhibiting memory [135].
γδ T Cells in Skin Disease
γδ T Cells in Skin Infections
γδ T cells by themselves are generally insufficient for sterilising immunity. However, Tcrb −/− x Tcrd −/− mice are more susceptible to viral infection indicating that these cells do have a protective role [19, 150]. Different subsets of γδ T cells may be mobilised in temporally and geographically distinct manner for early containment of damage, or later in the response for tissue repair and wound healing [18, 19]. In various models of cutaneous infection and vaccination, γδ T cells are recruited early and produce cytokines, often before the development of an αβ T cell response [100, 125]. The protozoan Leishmania is transmitted to its mammalian host through sandfly bite, and γδ T cell numbers are elevated in the blood of patients presenting cutaneous, mucosal or visceral leishmaniasis [122]. In mice, subcutaneous Leishmania major infection leads to a systemic expansion of Vδ4+ γδ T cells [121]. Antibody-mediated depletion of γδ T cells culminates in larger cutaneous lesions with higher number of parasites [120]. In Tcrα −/− mice, γδ T cells are instrumental in limiting pathogen spread and pathogen-associated damage, as shown in mouse models of footpad and corneal infection with Herpes simplex virus-1 (HSV-1). Mice with double deficiency of αβ and γδ T cells have larger epithelial lesions, higher viral load and dissemination, and succumb to lethal viral encephalitis [125]. In cutaneous Staphylococcus aureus infection, mice with γδ T deficiency have significantly larger skin lesions, higher bacterial load, and impaired recruitment of neutrophils to the infected site [23]. The ability of DETC to rapidly produce IL-17 in an IL-1, IL-23 and TLR2 dependent manner is critical for protection. In systemic S. aureus infection, a population of CD44+CD27− Vγ4+ γδ T cells that persists at the infection site and in draining lymph nodes after clearance of primary infection is found to rapidly expand and produce high amount of IL-17 upon secondary infection, displaying features of memory [98].
In addition, γδ T cells may modulate and shape the downstream adaptive response in a multitude of different ways in different models of infection. Depletion of γδ T cells in calves reduces M. bovis-specific IgG2 and IFN-γ, and increases IL-4 production [91]. The absence of γδ T cells selectively reduces IgA but not IgG or IgM production elicited by oral immunisation with tetanus toxoid plus cholera toxin [36]. In Chagas disease patients, the presence of IL-10 producing CD4−CD8− γδ T cells in the blood correlates positively with improved clinical parameters of cardiac function [148].
γδ T Cells in Tissue Homeostasis and Wound Healing
In the gut and the skin, intraepithelial γδ T cells do not only have a role in infection and inflammation, but are also indispensable in tissue homeostasis and repair. In the mouse epidermis, DETCs are in constant contact with adjacent keratinocytes as well as Langerhans cells. At steady state, DETC is the primary population of constitutive Insulin-Like Growth Factor-1 (IGF-1) producers supporting the survival, albeit not the proliferation, of keratinocytes [65]. Deficiency of γδ T cells is associated with increased epithelial apoptosis [126].
Wound healing involves coordinated phases of inflammation, proliferation, reepithelialisation, deposition of extracellular matrix and tissue remodelling. During this process, keratinocytes rapidly migrate to and colonise the wound border, produce antimicrobial factors to prevent microbial invasion, and proliferate to re-epithelialise the wound. It is known that γδ T cell deficient mice have a wound healing defect, which can be restored with wildtype DETC [66, 155]. A recent study identified a RORγt+ IL-17A producing subset of DETC to have a role in wound healing [87]. IL-17 blockade delays wound closure in wildtype mice, and conversely administration of exogenous IL-17 or transfer of IL-17-sufficient γδ T cells restore wound healing in IL-17A−/− mice. IL-17 induces downstream production of a multitude of anti-microbial factors with barrier protective functions [79, 87].
In human skin, both αβ and γδ T cells are capable of producing IGF-1. Elevation of IGF-1 is seen in acute but not chronic wounds [136]. Intraepithelial γδ T cells of the skin and the intestine, but not intraepithelial αβ T cells nor lymphoid γδ T cells, are capable of Keratinocyte Growth Factor-1 (KGF-1) production [9]. Wound healing is delayed in the presence of a dominant negative KGF receptor mutant transgene [153]. Although there is no cutaneous wound healing defect in KGF-1 deficient mice due to functional compensation by KGF-2 in the skin, exogenous KGF-1 rescues the wound healing defect of Tcrd −/− skin organ culture in vitro [66]. In normal wound healing, DETC-derived KGF-1 induces hyaluronan production from keratinocytes, which has a role in recruiting macrophages to the site [67]. Consequentially macrophage infiltration is defective in Tcrd −/− mice.
In mice but not humans, wound repair is accompanied by hair follicle regeneration, a process known as wound-induced hair neogenesis (WIHN). This process is dependent on the production of FGF-9 by dermal γδ T cells [39].
γδ T Cells in Psoriasis
The role of the IL23-IL17 axis in psoriasis pathogenesis is well recognised. IL-17A, IL-17 F, IL-22 and IL-21 are found to be elevated in the skin and blood of psoriatic patients, and IL-23 is selectively elevated in psoriatic lesions compared to non-lesional skin. Intradermal injection of IL-21 or IL-23 causes epithelial hyperplasia or acanthosis, one of the hallmarks of human psoriasis. IL-17A also upregulates the expression of keratin 17, a classical psoriatic disease marker, on keratinocytes [15]. IL-23R polymorphism is implicated in the pathogenesis of psoriasis [17]. While earlier studies have focused on Th17 cells, it is now known that innate cells including γδ T cells [16] and NKT cells [26] are major producers of IL-17 in psoriatic skin lesions.
In murine models of spontaneous psoriasitic dermatitis, disease progression correlates with a loss of Vγ5+ DETC and infiltration of IL-17+ dermal γδ T cells [4, 38]. There is compelling evidence indicating a pathogenic role of IL-17 producing dermal γδ T cells in various models of spontaneous and induced psoriatic dermatitis in mice [15, 16, 84, 86, 101, 140]. These models include topical application of imiquimod cream, a TLR7 agonist, or intradermal injection of IL-23 . IL-17A, IL-17 F and IL-22 deficient mice are protected from psoriatic dermatitis indicating the pathogenic role of these cytokines [101, 140]. Moreover, infiltrating dermal γδ T cells and RORγt+ ILC, rather than Th17 cells, have been shown to be the primary IL-17 producers, and are necessary and sufficient for psoriatic pathogenesis. More recently, Gray et al. found SOX13-dependent Vγ4+ Tγδ17 cells to be the specific pathogenic population in their model of psoriatic dermatitis [45]. These cells proliferate in skin draining lymph nodes and home to the inflamed skin. In a CD45.1+ C57BL/6 substrain, which carries a Sox13 mutation, the neonatal development of these Vγ4+ T cells is impaired, and these mice are protected from psoriasis development.
In human psoriasis patients, dermal IL-17-producing γδ T cells are found to be increased in psoriatic lesions [16]. Specifically, a population of CLA+CCR6+ Vγ9+Vδ2+ cells is reduced in the peripheral blood and increased in psoriatic skin lesions suggesting disease-associated redistribution of these cells. These cells produce IL-17A, and are capable of activating keratinocytes in vitro in a TNF-α and IFN-γ dependent manner [78].
There have been much recent efforts geared towards targeting the IL-23/IL-17 axis in the clinic. While targeting the p40 chain of IL-23/IL-12 is potentially risky due to its pleiotropic effect, the anti-IL-17 antibodies AIN457 (secukinumab) and LY2439821 (ixekizumab), and anti-IL-17R antibody AMG 827 (brodalumab) have shown promising efficacy in phase II clinical trials involving cases of chronic psoriatic plaques [15, 60, 81, 102].
Clinical Aspects of γδ T Cell Biology
Focus on Human Vγ9+Vδ2+ Cells as an Example
Vγ9+Vδ2+ cells constitute the major γδ T population in the peripheral blood of healthy humans. These cells are uniquely able to recognise self- and microbial- derived phosphoantigens in a TCR-dependent, MHC-unrestricted manner. This TCR binds isopentenyl pyrophosphate, an isoprenoid intermediate of the human mevalaonate pathway, and also the microbial isoprenoid (HMBPP), an intermediate of the 2-C-methyl-D-erythritol-4 phosphate (MEP) pathway for isoprenoid biosynthesis [96]. This pathway is shared by a broad spectrum of prokaryotic and eukaryotic pathogens. The ability of these cells to bind an evolutionarily conserved group of invariant molecules via the TCR is consistent with their purported role as early rapid effectors. It has been shown that phosphoantigen recognition requires the presence of APC but is independent of MHC, MR1 or CD1. The nature of presentation of phosphoantigen remains to be resolved. The mitochondrial protein F1 ATPase (F1 adenosine triphosphatase) is a candidate phospho-antigen presenting molecule, but its involvement remains to be proven. Recent studies provided compelling evidence for an antigen-presenting role of BTN3A1, a butyrophilin-like extended B7 family member, in the binding of phospho-antigen to the Vγ9Vδ2 TCR [123, 145].
Vγ9+Vδ2+ T cells may have promising clinical potential especially in anti-tumour therapy [44, 96, 147, 158]. These cells are able to recognise a broad range of tumour cells, and have been demonstrated to be cytolytic towards a variety of tumour cells in vitro. Interestingly, transformed cells have been shown to upregulate the mevalonate pathway and accumulate IPP intracellularly, pointing to possibilities of ex vivo expansion of these γδ T cells and targeting a conserved pathway of tumour cell metabolism. The identification of the minimal binding moiety for γδ TCR, which is a five-carbon alkenyl chain with a pyrophosphate moiety, facilitates the mining and synthesis of phosphoantigen-based agonists for ex vivo manipulation and expansion of these cells.
In mouse models γδ T cells have been shown to mediate anti-tumour immunity via rapid early production of cytokines [37, 85]. In a murine sarcoma model, chemotherapy induces an early rapid infiltration of Tγδ17 cells into the tumour bed that is required for the subsequent infiltration of CD8+ CTL and anti-tumour chemotherapeutic efficacy [85].
Tools for Probing γδ T Cells
The inherently complex biology of γδ T cells has proven to be challenging in efforts to elucidate their behaviour and functions. In recent years, the combination of several different tools has revealed previously unappreciated aspects of their biology. The development of tetramer-staining reagents has provided new information on the role of TCR ligation in generation of γδ T cells [68, 75]. Further structural studies are likely to yield useful novel insights on the mechanism by which γδ TCR recognise a diverse range of antigens.
The vast majority of γδ T cells in mice and humans are tissue resident cells, which pose a particular challenge to flow cytometry-based studies that require tissue disintegration and single cell isolation. Enzymatic digestion has to be sufficient for single cell isolation on one hand and preserve important yet enzyme-sensitive surface molecules on the other. The availability of genetically fluorescent tagged mice and powerful intravital imaging platforms have solved some of these issues, offering the opportunity to study the cells in situ in real-time [20, 32, 47, 132]. The discovery that the skin contains two segregated γδ populations with distinct biology in the epidermis and the dermis is a testimony of the power of the combination of such technologies. More recently, immunological synapses between keratinocytes and DETC have been characterised using intravital dynamics-immunosignal correlative microscopy [24].
Summary
γδ T cells have unique features that make them highly relevant to immune surveillance in the host, and not a vestigial population from the evolutionary perspective. These cells recognise self-derived stress signals as well as microbial invariant molecular patterns without the need for antigen processing or presentation in an MHC restricted manner. They are localised and self-renew at peripheral sites, thus increase the probability of detecting antigens.
Some γδ T populations express invariant or oligoclonal TCR. Although not a universal feature of γδ T cells, in combination with a restricted localization, this allows the cells to respond rapidly and at an effective magnitude without having to clonally expand, with a TCR specificity evolutionarily selected to recognise antigens likely to be encountered at peripheral sites. γδ T cells are often present in a semi-activated state, allowing them to rapidly acquire effector function without TCR ligation. The myriad interactions between γδ T cells and other cell types allow them to polarise downstream responses to facilitate an efficacious immunological outcome.
Appendix
*There are different nomenclatures for the different γδ TCR V genes, namely the Tonegawa & Heilig, Garman and Hayday systems. For example Vγ5 in the Tonegawa’s system corresponds to Vγ3 in Garman’s and GV1S1 in the Hayday’s system. The Heilig & Tonegawa system is used in this manuscript, and in the International Immunogenetics Information System (IMGT).




Learning Objectives
Related posts:




Stay updated, free articles. Join our Telegram channel
Full access? Get Clinical Tree
