Buruli ulcer (BU), caused by the environmental organism Mycobacterium ulcerans and characterized by necrotizing skin and bone lesions, poses important public health issues as the third most common mycobacterial infection in humans. Pathogenesis of M ulcerans is mediated by mycolactone, a necrotizing immunosuppressive toxin. First-line therapy for BU is rifampin plus streptomycin, sometimes with surgery. New insights into the pathogenesis of BU should improve control strategies.
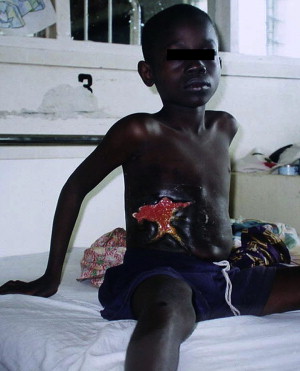
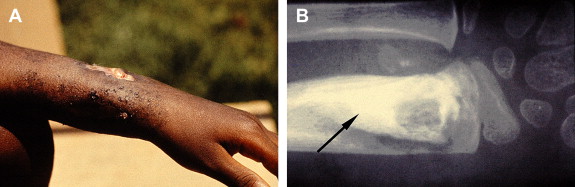
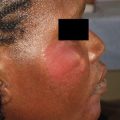
Buruli ulcer (BU), the third most common mycobacterial infection in humans next to tuberculosis and leprosy, is an emerging infection caused by Mycobacterium ulcerans . BU is characterized by indolent, typically painless necrotizing skin lesions ( Figs. 1 and 2 A). Approximately 10% of patients develop bone involvement subjacent to skin lesions or metastatic osteomyelitis from lymphohematogenous spread of M ulcerans (see Fig. 2 B). Pathogenesis is mediated by mycolactone, a diffusible, necrotizing, immunosuppressive, polyketide-derived macrolide toxin secreted by M ulcerans . In 1962, the disease was named after Buruli County, Uganda, now called Nakasongola District, where the epidemic was documented first. Other names include Bairnsdale, Kakerifu, Kasongo, or Searls’ ulcer.
Epidemiology
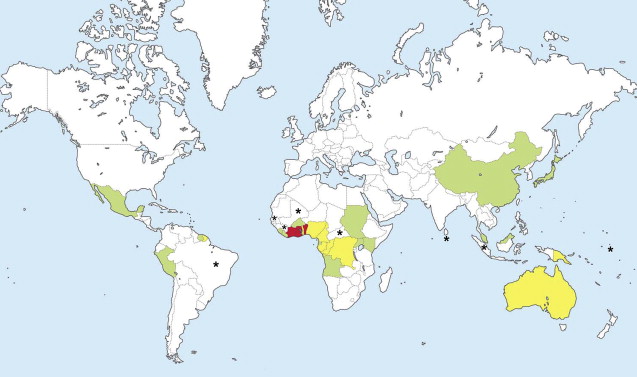
In 1998, the World Health Organization (WHO) recognized BU as a reemerging infectious disease in West and Central Africa, with a significant public health impact. The reported incidence rates of BU are highest in West Africa, especially Benin, Ghana, and Côte d’Ivoire. However, BU is reported in about 30 countries ( Fig. 3 ), and growing evidence suggests that BU is more widespread than earlier thought. BU prevails in rural tropical wetlands, especially areas with stagnant water, including ponds and swamps. However, BU is also acquired without wetland exposure.
The rapid reemergence of BU, beginning in the early 1980s, particularly in areas where people are engaged in manual agriculture in wetlands, may be attributable to the man-made alterations to the environment, such as deforestation and other topographic alterations, which increase the amount of wetlands. Changes in global temperature and precipitation patterns further promote the reemergence of BU.
The WHO reports indicate that more than 5000 people are diagnosed with BU annually, but many cases are undiagnosed because of the geopolitical and socioeconomic factors in endemic countries. Children (5–15 years old) have the highest incidence of BU, with most lesions on the lower extremities. BU is a growing public health problem, with psychosocial and socioeconomic implications in endemic regions. Up to 60% of patients with BU suffer from disabling and stigmatizing sequelae, including scarring, contractures, and bone destruction. Minimizing disability through treatment, both antimicrobial and surgical, and physiotherapy is, therefore, important in BU management. Imported BU is occasionally diagnosed in the United States, Canada, and Europe.
BU is directly related to environmental factors and thus considered noncontagious. The most possible mode of transmission is local, minor, often unnoticed skin trauma that permits inoculation of M ulcerans . The estimated incubation period is 2 to 3 months. Because M ulcerans DNA is detectable in some aquatic insects, the role of insects as vectors that infect humans by biting is under investigation. In Australia, some investigators propose that BU is a zoonosis transmitted by mosquitoes from indigenous marsupials (eg, possums and koalas) to humans. M ulcerans DNA was found in mosquitoes during an outbreak of BU in humans in Australia, and the seasonal incidence of BU in humans correlates with that of notifiable arthropod-borne diseases in Victoria. In Africa, terrestrial mammals are being investigated as reservoirs of M ulcerans .
Risk factors for BU within endemic areas include failure to wear protective clothing, exposure to unprotected natural water sources, and inadequate care of minor skin wounds. Human immunodeficiency virus seropositivity may increase the risk for BU or be associated with aggressive BU.
BCG vaccination has some effect on BU. Several reports suggest that BCG vaccination provides some protection against BU, for 6 to 12 months after vaccination, and that neonatal BCG vaccination reduces the risk of BU osteomyelitis in those who acquire BU as children or adults. However, a case-control study concluded that BCG vaccination is not protective against BU. Prophylactic and therapeutic vaccines based on DNA engineering and virulence factors, including mycolactone, are under study (BuruliVac Project). Intravenous immunoglobulin to neutralize mycolactone is not available.
Microbiology of M ulcerans
Standard and real-time polymerase chain reaction (PCR) techniques have been used to identify M ulcerans , primarily by detecting 2 M ulcerans insertion sequences (IS 2404 and IS 2606 ), in the environment in Australia and West Africa. Improved M ulcerans DNA extraction procedures enhance environmental detection, thereby advancing the understanding of reservoirs. Portaels and colleagues reported the first direct isolation of M ulcerans from nature in 2008 from a water strider, an aquatic insect that does not bite humans.
Unlike M leprae and M tuberculosis (the pathogens for leprosy and tuberculosis, respectively), M ulcerans produces a necrotizing, immunosuppressive, polyketide-derived macrolide toxin, called mycolactone. Genes in a virulence plasmid of M ulcerans , controlled by SigA-like promoters, encode for the synthesis of mycolactone. Identification of SigA-like promoters led to the development of M ulcerans –green fluorescent protein. This tagged protein linking fluorescence with toxin gene expression is a potential tool for studying BU pathogenesis and transmission.
M ulcerans shares some environmental, molecular, and clinical features with M marinum , a water-associated organism that causes granulomatous skin lesions in humans, often called “swimming pool” or “fish tank” granuloma. Comparative genomics indicate that M ulcerans likely diverged from M marinum , acquiring a 174-kb virulence plasmid (pMUM001) with genes coding for mycolactone production and 10 proteins, all potential targets for vaccine development or serodiagnosis. Accordingly, phenolic mycosides of M ulcerans and M marinum are identical, and sequences for the 16S ribosomal RNA (rRNA) gene are nearly identical. As M ulcerans evolved toward becoming an intracellular organism, like M marinum , nonessential genes were lost, which may have increased the pathogenicity.
Gene sequences of the 3′ end of the 16S rRNA of M ulcerans vary by geographic origin, dividing M ulcerans broadly into African, American, Asian, and Australian strains, with many substrains on each continent. Each major strain generally differs in clinical presentation, mycolactone type and virulence, and host immune responses. Mycolactone type coding by geographic origin includes A/B (Africa, the most pathogenic), C (Asia, Australia), and D (Asia).
Molecular genetic techniques are slowly unraveling the evolution of M ulcerans . M ulcerans isolates from localized foci within endemic regions often show a high degree of genomic similarity (ie, clonal populations) with a lack of insertional-deletional genomic polymorphisms, underscoring a requirement for single-nucleotide polymorphism (SNP) analysis to differentiate substrains of M ulcerans within those areas. Identifying SNPs and establishing SNP typing assays are increasingly defining the microepidemiology, genetic diversity, and evolution of M ulcerans . For example, SNP analyses of M ulcerans in Ghana differentiate 54 M ulcerans strains into 13 SNP haplotypes, yet a geographically focal transmission.
Microbiology of M ulcerans
Standard and real-time polymerase chain reaction (PCR) techniques have been used to identify M ulcerans , primarily by detecting 2 M ulcerans insertion sequences (IS 2404 and IS 2606 ), in the environment in Australia and West Africa. Improved M ulcerans DNA extraction procedures enhance environmental detection, thereby advancing the understanding of reservoirs. Portaels and colleagues reported the first direct isolation of M ulcerans from nature in 2008 from a water strider, an aquatic insect that does not bite humans.
Unlike M leprae and M tuberculosis (the pathogens for leprosy and tuberculosis, respectively), M ulcerans produces a necrotizing, immunosuppressive, polyketide-derived macrolide toxin, called mycolactone. Genes in a virulence plasmid of M ulcerans , controlled by SigA-like promoters, encode for the synthesis of mycolactone. Identification of SigA-like promoters led to the development of M ulcerans –green fluorescent protein. This tagged protein linking fluorescence with toxin gene expression is a potential tool for studying BU pathogenesis and transmission.
M ulcerans shares some environmental, molecular, and clinical features with M marinum , a water-associated organism that causes granulomatous skin lesions in humans, often called “swimming pool” or “fish tank” granuloma. Comparative genomics indicate that M ulcerans likely diverged from M marinum , acquiring a 174-kb virulence plasmid (pMUM001) with genes coding for mycolactone production and 10 proteins, all potential targets for vaccine development or serodiagnosis. Accordingly, phenolic mycosides of M ulcerans and M marinum are identical, and sequences for the 16S ribosomal RNA (rRNA) gene are nearly identical. As M ulcerans evolved toward becoming an intracellular organism, like M marinum , nonessential genes were lost, which may have increased the pathogenicity.
Gene sequences of the 3′ end of the 16S rRNA of M ulcerans vary by geographic origin, dividing M ulcerans broadly into African, American, Asian, and Australian strains, with many substrains on each continent. Each major strain generally differs in clinical presentation, mycolactone type and virulence, and host immune responses. Mycolactone type coding by geographic origin includes A/B (Africa, the most pathogenic), C (Asia, Australia), and D (Asia).
Molecular genetic techniques are slowly unraveling the evolution of M ulcerans . M ulcerans isolates from localized foci within endemic regions often show a high degree of genomic similarity (ie, clonal populations) with a lack of insertional-deletional genomic polymorphisms, underscoring a requirement for single-nucleotide polymorphism (SNP) analysis to differentiate substrains of M ulcerans within those areas. Identifying SNPs and establishing SNP typing assays are increasingly defining the microepidemiology, genetic diversity, and evolution of M ulcerans . For example, SNP analyses of M ulcerans in Ghana differentiate 54 M ulcerans strains into 13 SNP haplotypes, yet a geographically focal transmission.
Pathogenesis and immunity
Initial infection is primarily related to 2 properties of M ulcerans : optimal growth at temperatures (30°C–33°C) slightly below the core body temperature and production of mycolactone. The temperature requirement of M ulcerans favors the development of lesions in cooler tissues, especially the skin and subcutaneous tissue. Mycolactone destroys tissues by apoptosis and necrosis ( Fig. 4 ) and suppresses host immune responses.
Related posts:
 Outbreak of Nontuberculous Mycobacterial Disease in the Central Pacific
Outbreak of Nontuberculous Mycobacterial Disease in the Central Pacific
 Dermatology Aboard the USNS COMFORT: Disaster Relief Operations in Haiti After the 2010 Earthquake
Dermatology Aboard the USNS COMFORT: Disaster Relief Operations in Haiti After the 2010 Earthquake
 Chagas Disease: Coming to a Place Near You
Dermatology in Botswana: The American Academy of Dermatology’s Resident International Grant
Chagas Disease: Coming to a Place Near You
Dermatology in Botswana: The American Academy of Dermatology’s Resident International Grant
 Widespread Use of Toxic Skin Lightening Compounds: Medical and Psychosocial Aspects
Human Immunodeficiency Virus and Leprosy: An Update
Widespread Use of Toxic Skin Lightening Compounds: Medical and Psychosocial Aspects
Human Immunodeficiency Virus and Leprosy: An Update
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


