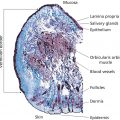
Francis McGlone1, David Reilly2, and Zoe Diana Draelos3 1 School of Natural Sciences & Psychology, Liverpool John Moores University, Liverpool, UK 2 Unilever Research, Colworth Science Park, Bedford, UK 3 Dermatology Consulting Services, PLLC, High Point, NC, USA The primary sensory modality subserving the body senses is collectively described as the somatosensory system and comprises all those peripheral afferent nerve fibers, and specialized receptors, subserving cutaneous, and proprioceptive sensitivity. The latter processes information about limb position and muscle forces which the central nervous system uses to monitor and control limb movements and, via elegant feedback and feedforward mechanisms, ensure that a planned action or movement is executed fluently. This chapter focuses on sensory inputs arising from the skin surface – cutaneous sensibility – and describes the neurobiological processes that enable the skin to “sense.” Skin sensations are multimodal and are classically described as sensing the four submodalities of touch, temperature, itch, and pain. We also consider the growing evidence for a fifth submodality, present only in hairy skin, which is preferentially activated by slowly moving, low force, mechanical stimuli. This brief introduction to somatosensation starts with the discriminative touch system. Sensation enters the peripheral nervous system via sensory axons that have their cell bodies sitting just outside the spinal cord in the dorsal root ganglia, with one ganglion for each spinal nerve root. Neurons are the building blocks of the nervous system and somatosensory neurons are unique in that, unlike most neurons, the electrical signal does not pass through the cell body but the cell body sits off to one side, without dendrites. The signal passes directly from the distal axon process to the proximal process which enters the dorsal half of the spinal cord, and immediately turns up the spinal cord forming a white matter column, the dorsal columns, which relay information to the first brain relay nucleus in the medulla. These axons are called the primary afferents, because they are the same axons that carry the signal into the spinal cord. Sensory input from the face does not enter the spinal cord but instead enters the brainstem via the trigeminal nerve (one of the cranial nerves). Just as with inputs from the body, there are four modalities of touch, temperature, itch, and pain, with each modality having different receptors traveling along different tracts projecting to different targets in the brainstem. Once the pathways synapse in the brainstem, they join those from the body on their way up to a relay in the thalamus and then on to higher cortical structures. Sensory information arising from the skin is represented in the brain in the primary and secondary somatosensory cortex, where the contralateral body surfaces are mapped in each hemisphere. The skin is the most extensive and versatile organ of the body and in a fully grown adult covers a surface area approaching 2 m2. This surface is far more than just a passive barrier. It contains in excess of two million sweat glands and five million hairs covering all surfaces, apart from the soles of the feet and the palms of the hands (glabrous skin). Evidence is also emerging that nonglabrous skin contains a system of nerves that code specifically for the pleasant properties of touch. Skin consists of an outer, waterproof, stratified squamous epithelium of ectodermal origin – the epidermis – plus an inner, thicker, supporting layer of connective tissue of mesodermal origin – the dermis. The thickness of this layer varies from 0.5 mm over the eyelid to >5.0 mm over the palm and sole of the foot. Of the four “classic” submodalities of the somatosensory system, discriminative touch subserves the perception of pressure, vibration, and texture and relies upon four different receptors in the digit skin: These are collectively known as low threshold mechanoreceptors (LTMs), a class of cutaneous receptors that are specialized to transduce mechanical forces impinging the skin into nerve impulses. The first two are classified as fast adapting (FA) as they only respond to the initial and final contact of a mechanical stimulus on the skin, and the second two are classified as slowly adapting (SA) as they continue firing during a constant mechanical stimulus. A further classification relates to the LTM’s receptive field (RF; i.e. the surface area of skin to which they are sensitive). The RF is determined by the LTM’s anatomic location within the skin, with those near the surface at the dermal–epidermal boundary, Meissner corpuscles, and Merkel disks, having small RFs, and those lying deeper within the dermis, Pacinian corpuscles (), and Ruffini endings, having large RFs (Figure 4.1). Psychophysical procedures have been traditionally employed to study the sense of touch where differing frequencies of vibrotactile stimulation are used to quantify the response properties of this sensory system. Von Bekesy [1] was the first to use vibratory stimuli as an extension of his research interests in audition. In a typical experiment, participants were asked to respond with a simple button‐press when they could just detect the presence of a vibration presented to a digit, within one of two time periods. This two‐alternative force choice (2‐AFC) paradigm provides a threshold‐tuning curve, the slopes of which provide information about a particular class of LTM’s response properties. Bolanowski et al. [2] proposed that there are four distinct psychophysical channels mediating tactile perception in the glabrous skin of the hand. Each psychophysically determined channel is represented by one of the four anatomic end organs and nerve fiber subtypes, with frequencies in the 40–500 Hz range providing a sense of “vibration,” transmitted by PC (PC channel or FAI); Meissner corpuscles being responsible for the sense of “flutter” in the 2–40 Hz range (NPI channel or FAII); the sense of “pressure” being mediated by Merkel disks in the 0.4–2.0 Hz range (NPIII or SAI); and Ruffini end organs producing a “buzzing” sensation in the 100–500 Hz range (NPII or SAII). Neurophysiologic studies support this model, but there is still some way to go to link the anatomy with perception (Table 4.1). There have been relatively few studies of tactile sensitivity on hairy skin, the cat being the animal of choice for most of these studies. Mechanoreceptive afferents (Aβ fibers) have been described that are analogous to those found in human glabrous skin (FAI, FAII, SAI, SAII), and Essick and Edin [3] have described sensory fibers with these properties in human facial skin. The relationship between these sensory fibers and tactile perception is still uncertain. Sensory axons are classified according to their degree of myelination, the fatty sheath that surrounds the nerve fiber. The degree of myelination determines the speed with which the axon can conduct nerve impulses and hence the nerves conduction velocity. The largest and fastest axons are called Aα and include some of the proprioceptive neurons, such as the muscle stretch receptors. The second largest group, called Aβ, includes all of the discriminative touch receptors being described here. Pain, itch, and temperature include the third fourth and fifth groups, Aδ and C‐fibers. Electrophysiological studies on single peripheral nerve fibers innervating the human hand have provided a generally accepted model of touch that relates the four anatomically defined types of cutaneous or subcutaneous sense organs to their neural response patterns [4]. The technique used in these studies is called microneurography and involves inserting a fine tungsten microelectrode, tip diameter <5 μm, through the skin and into the underlying median nerve which innervates the thumb and first two digits (Figure 4.2). The cutaneous somatosensory system detects changes in ambient temperature over an impressive range, initiated when thermal stimuli that differ from a homeostatic set‐point excite temperature‐specific sensory nerves in the skin and relay this information to the spinal cord and brain. It is important to recognize that these nerves code for temperature change, not absolute temperature, as a thermometer does. The system does not have specialized receptor end organs such as those found with LTMs but uses free nerve endings throughout skin to sense changes in temperature. Within the innocuous thermal sensing range, there are two populations of thermosensory fibers, one that responds to warmth (warm receptors) and one that responds to cold (cold receptors), and include fibers from the Aδ and C range. Specific cutaneous cold and warm receptors have been defined as slowly conducting units that exhibit a steady‐state discharge at constant skin temperature and a dynamic response to temperature changes [6, 7]. Cold‐specific and warm‐specific receptors can be distinguished from nociceptors that respond to noxious low and high temperatures <20 °C and >45 °C) [8, 9], and also from thermosensitive mechanoreceptors [6, 10]. Standard medical textbooks describe the cutaneous cold sense in humans as being mediated by myelinated A‐fibers with CVs in the range 12–30/ms [11], but recent work concludes that either human cold‐specific afferent fibers are incompletely myelinated “BC” fibers, or else there are C as well as A cold fibers, with the C‐fiber group contributing little to sensation (Figure 4.3) [12]. Figure 4.1 A cross‐sectional perspective of (a) glabrous and (b) hairy skin. (Source: R.T. Verrillo, artist. Reproduced with permission.) Table 4.1 Main characteristics of primary sensory afferents innervating human skin. The free nerve endings for cold‐sensitive or warm‐sensitive nerve fibers are located just beneath the skin surface. The terminals of an individual temperature‐sensitive fiber do not branch profusely or widely. Rather, the endings of each fiber form a small, discretely sensitive point, which is separate from the sensitive points of neighboring fibers. The total area of skin occupied by the receptor endings of a single temperature‐sensitive nerve fiber is relatively small (approximately 1 mm in diameter), with the density of these thermosensitive points varying in different body regions. In most areas of the body, there are 3–10 times as many cold‐sensitive points as warm‐sensitive points. It is well established from physiologic and psychologic testing that warm‐sensitive and cold‐sensitive fibers are distinctively different from one another in both structure and function.
CHAPTER 4
The Somatosensory System and Sensitive Skin
Introduction
Peripheral nervous system
Touch
Temperature

Class
Modality
Axonal diameter (μm)
Conduction velocity (m/s)
Myelinated
Aα
Proprioceptors from muscles and tendons
20
120
Aβ
Low threshold mechanoreceptors
10
80
Aδ
Cold, noxious, thermal
2.5
12
Unmyelinated
C‐pain
Noxious, heat, thermal
1
<1
C‐tactile
Light stroking, gentle touch
1
<1
C‐tutonomic
Autonomic, sweat glands, vasculature
1
<1
Pain
Related posts:


Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


