, Teresa S. Wright2, Crystal Y. Pourciau3 and Bruce R. Smoller4
(1)
Department of Pathology & Immunology, Baylor College of Medicine and Texas Children’s Hospital, Houston, Texas, USA
(2)
Departments of Dermatology and Pediatrics, University of Tennessee Health Science Center, Memphis, TN, USA
(3)
Departments of Dermatology and Pediatrics, Baylor College of Medicine and Texas Children’s Hospital, Houston, Texas, USA
(4)
Department of Pathology and Laboratory Medicine, University of Rochester School of Medicine and Dentistry, Rochester, NY, USA
With special contributions from Arshad Al Mashhadani, MD, Baylor College of Medicine.
12.1 Herpes Simplex Virus
12.1.1 Clinical Features
In the United States, approximately 1,500 cases of neonatal herpes simplex virus infection are diagnosed annually [1]. The clinical manifestations of neonatal herpes are classified in three categories: SEM disease (limited to the skin, eyes and mouth), central nervous system disease, and disseminated disease. Skin lesions may or may not be present in central nervous system and disseminated infections. Approximately 60–70 % of infants with central nervous system disease have associated skin vesicles and erythematous erosions. Similarly, most of those with disseminated disease have cutaneous manifestations, with only 20 % of infants not having skin vesicles at any time during their illness.
The majority of oro-labial herpetic disease is caused by herpes simplex virus-1 (HSV-1). Approximately 70–80 % of adults are HSV-1 seropositive by age 40 [2]. Genital herpes infections can be caused by HSV-1 or HSV-2. Historically, the majority of genital infections are caused by HSV-2. However, there has been an increase in the incidence of genital infections by HSV-1.
Classic lesions of cutaneous herpes infection consist of small grouped monomorphous vesicles on an erythematous background (Fig. 12.1). These evolve into shallow erosions and crusted papules, which typically heal without leaving permanent scarring.
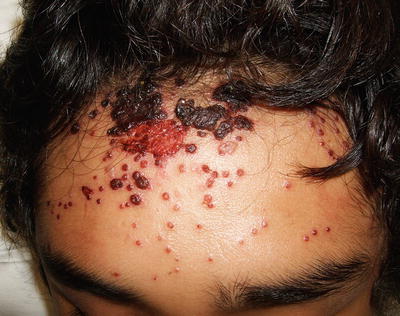
Fig. 12.1
Herpes simplex virus lesions present as multiple monomorphic, erythematous vesicles, some of which coalesce to form large erosions with scalloped borders and overlying hemorrhagic crust
12.1.2 Histology
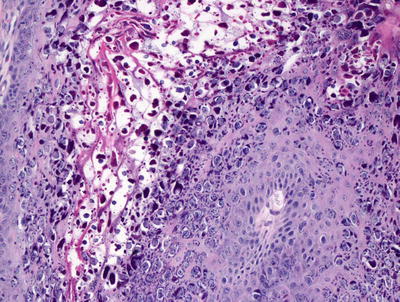
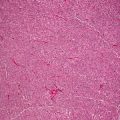
Skin biopsy of mucocutaneous HSV infection primarily demonstrates intraepidermal changes. Keratinocytes exhibit balloon cell changes and reticular degeneration, resulting in acantholysis and suprabasilar blister formation (Fig. 12.2). Blisters appear at multiple levels of the epidermis, and can be variable in size. Intranuclear inclusions are present within keratinocytes. These appear as clear nuclei with chromatin margination (Fig. 12.3). Molding of keratinocyte nuclei and multinucleation are also present. Follicular herpetic infection demonstrating the same changes within follicular keratinocytes has been described more commonly in immunosuppressed patients [3, 4]. Verrucous lesions with marked epidermal hyperplasia have also been associated with underlying immunosuppression [5].
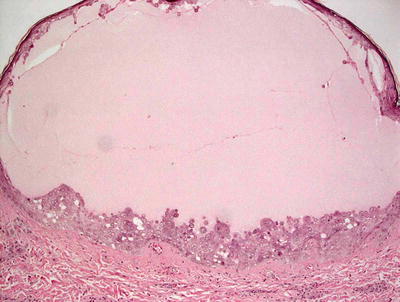
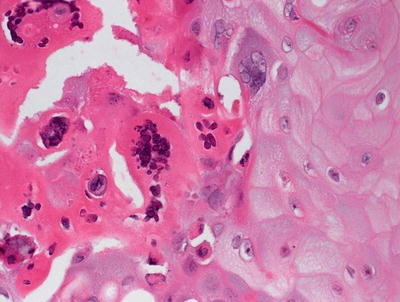
Fig. 12.2
Herpes simplex virus infection demonstrates an intraepidermal blister with abundant nuclear viral inclusions
Fig. 12.3
Multinucleated cells with nuclear clearing, molding, and acantholysis characterize viral cytopathic changes in herpes simplex virus infection
The underlying dermis frequently demonstrates a marked inflammatory infiltrate, especially in primary infections. The infiltrate is polymorphous and contains many neutrophils, resembling leukocytoclastic vasculitis in some cases. Inflammation of eccrine ducts is also encountered in cutaneous herpes infection [6]. A variety of atypical reactions have been described within the dermis, including marked lymphoid atypia, plasmacellular atypia, presence of CD30-positive cells, florid pseudolymphomatous infiltrate, and granulomatous reaction pattern [7–11]. As a rule, reinfections are less inflammatory than primary infections with herpes simplex virus, although it is difficult to make this distinction with any degree of accuracy .
12.1.3 Pathogenesis
Herpes simplex virus is a DNA virus in the alpha herpes virus family. HSV exists as two distinct types: herpes simplex virus-1 (HSV-1) and herpes simplex virus-2 (HSV-2) [12]. HSV-1 mainly causes oral lesions, and HSV-2 commonly causes genital herpetic ulcers and neonatal herpes [13–15]. The DNA genomes of HSV-1 and HSV-2 share approximately 50 % homology with significant antigenic cross-reactivity [16]. HSV-1 and HSV-2 can be distinguished based on glycoprotein G (gG-1 for HSV-1, and gG-2 for HSV-2) with an antigen-specific antibody response, as well as by molecular tests, including DNA sequencing and real-time polymerase chain reaction (PCR) [17, 18]. HSV-2 is transmitted sexually with persistent chronic infection. HSV-2 infects epithelial cells and nerve endings with retrograde axonal transport to establish chronic infection in the sacral ganglia. The virus can lie dormant and remain “latent” in the ganglia until reactivation by a triggering event, in which the virus travels along the nerve axon to the skin and mucosal surfaces [15].
The innate immune response is the first line of host defense that limits HSV spread, and it is important in the activation of the adaptive immune response [19]. Toll-like receptors (TLRs) are important in the recognition of HSV antigens by host immune cells. Viral glycoproteins on HSV virions are first detected by TLR2 present on the cell surface [20, 21]. Viral replication and synthesis of viral intermediate double-stranded DNA are sensed by endosomal TLR3 [19, 22]. Viral double-stranded DNA is recognized by TLR9, and consequently stimulates interferon-α production, and the induction of interferon-response genes that encode antiviral effectors to limit viral replication and spread [23–25].
Cellular apoptosis and necroptosis are other important host defense mechanisms in the elimination of pathogen-infected cells. Necroptosis is a regulated form of necrosis that can occur in the absence of caspase activity [26]. Since the death of HSV-infected cells can limit viral replication and spread, HSV has evolved ways to evade cellular apoptosis. It has been found that HSV encodes anti-apoptotic viral proteins to block apoptosis. The immediate-early viral genes, such as ribonucleotide reductase large subunit (R1), are capable of inhibiting apoptosis and necroptosis during HSV infection [27]. Receptor-interacting kinase-3 (RIP3) has emerged as a central signaling molecule in necroptosis [28]. R1 is a suppressor of caspase-8-induced apoptosis and RIP3-induced necroptosis in human cells [29–31]. Thus, HSV appears to block cell death via the suppression of RIP3 signaling during HSV infection to allow viral replication in live host cells .
12.2 Human Herpesviruses 6, 7 and 8
12.2.1 Clinical Features
Human herpesviruses (HHV) are common in the general population with evidence of previous HHV-6 and HHV-7 infections present in nearly everyone over 6 years of age. It is estimated that 80–90 % of the entire population intermittently sheds these viruses in saliva. HHV-8 is somewhat less common than HHV-6 and HHV-7, with seroprevalence in the United States estimated at less than 10 % [32].
Infections with HHV-6, -7, and -8 are associated with a variety of cutaneous diseases. HHV-6 and HHV-7 are associated with the childhood eruption roseola infantum. The typical child with roseola is a toddler, who develops widespread morbilliform eruption after experiencing relatively high fever for about 3 days. HHV-6 and HHV-7 have also been implicated in pityriasis rosea, although a clear association has not been proven conclusively. Other conditions that may be associated with HHV-6 and HHV-7 infections include Stevens-Johnson syndrome, thrombocytopenic purpura, purpura fulminans, papular purpuric gloves and socks, Gianotti-Crosti syndrome, drug-induced hypersensitivity syndrome, and post-transplant graft vs. host disease. HHV-7 may also have a causative role in the development of lichen planus. HHV-8, also known as Kaposi sarcoma-associated herpes virus, has been found in endemic, classic, and post-transplant forms of the disease.
12.2.2 Histology
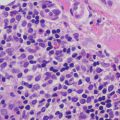
HHV-6 infection affects the skin, but lesions are rarely biopsied. The changes are often nonspecific, and include a superficial perivascular lymphocytic infiltrate. Scattered large and atypical cells with intranuclear and intracytoplasmic viral inclusions and cytopathic changes are seen in some cases [33]. Infected cells are described as medium- to large-sized lymphocytes with large centrally-located basophilic inclusions surrounded by halos [34].
The exact relationship between HHV-7 and skin infection is unknown. To date, there are no specific changes that are directly linked to HHV-7; however, there is emerging literature linking HHV-7 and pityriasis rosea [35–37]. The virus has also been linked to atypical lymphoid proliferations, although the data is less convincing [32]. No specific viral cytopathic changes in the skin that have been directly attributed to HHV-7.
Roseola infantum is caused by human herpesvirus, and demonstrates changes of spongiotic dermatitis, occasionally with vesicle formation. There is an underlying superficial perivascular lymphocytic infiltrate with papillary dermal edema and focal exocytosis [38].
While HHV-8 plays an important role in several cutaneous diseases, there are no specific viral cytopathic changes that are currently recognized [32]. HHV-8 has been intimately associated with Kaposi’s sarcoma, and it is readily identified in most cases on anti-HHV-8 immunostains [39]. Recently, HHV-8 has also been identified in eruptive cherry angiomas, but not in sporadic, commonly-occurring angiomas [40]. Rare cases of cutaneous lymphoma also demonstrate the presence of the virus [41, 42].
12.2.3 Pathogenesis
HHV-6 and HHV-7 are members of the Roseolovirus genus within the beta herpesvirinae subfamily [43]. They are genetically related to cytomegalovirus [44, 45]. HHV have a tropism for lymphocytes and macrophages, and have a long latency period [43]. The viruses can shut down DNA synthesis of infected host cells, induce protein synthesis and upregulate various cytokines and chemokines, causing viral cytopathic effects and ballooning appearance in affected cells [43].
HHV-8 is an oncogenic gamma herpesvirus [46]. It infects endothelial and epithelial cells, B lymphocytes, monocytes, and CD34+ hematopoietic progenitor stem cells [47]. It also targets antigen-presenting cells, including monocyte-derived dendritic cells [46]. HHV-8 has both latent and lytic cycles [48]. The virus is known to produce proteins that enable it to promote cell survival, evade the immune system, stimulate angiogenesis, and induce inflammation. Furthermore, it induces the production of vascular endothelial growth factor (VEGF) , which is a potent pro-angiogenic growth factor that induces vascular proliferation [47]. Through its latent gene expression, the virus is able to block endothelial cell apoptosis and promote cell survival, which may contribute to the pathogenesis of Kaposi’s sarcoma. HHV-8 also induces cytokine production that contributes to angiogenesis and inflammation characteristic of these vascular neoplasms [48].
12.3 Congenital Varicella Zoster Virus
12.3.1 Clinical Features
Intrauterine infection occurs in up to 25 % of women who contract primary varicella zoster virus during the first two trimesters of pregnancy. Approximately 12 % of infected babies will have congenital anomalies [49]. In pregnant women who are infected between the 8th and 26th weeks of pregnancy, about 2 % of fetuses will develop congenital varicella syndrome. This is characterized by cutaneous scarring, severe central nervous system damage, eye abnormalities, and limb deformities [5].
12.3.2 Histology
The histologic changes seen in congenital varicella zoster infection are largely the same as those seen in primary herpes simplex virus infections as described elsewhere in this chapter. At least one case has been reported in which the changes resemble the verrucous type of infection most commonly described in immunosuppressed patients [5, 51].
12.3.3 Pathogenesis
Congenital varicella is caused by infection with varicella zoster virus, which is a DNA virus of the herpesvirus family. Approximately 12–30 % of fetuses contract varicella transplacentally from mothers who contract the disease before the 24th week of pregnancy [51–53]. Congenital varicella syndrome with all of its manifestations occurs in less than 1 % of infants under the same conditions [54]. It is generally believed that as immunocompetence develops later in pregnancy, the fetus is able to mount an immune response to the invading virus, leading to a more benign disease course. The syndrome results from damage to the nervous system that occurs early in gestational development as the neurotropic virus invades these primitive cells [55]. As a result of incompletely developed immune system in fetuses, there is a very short latency period following viral infection of neural tissue. Upon reactivation, the viruses lead to maldevelopment of the musculoskeletal system as well as somatic and autonomic nervous system [56].
12.4 Varicella Zoster
12.4.1 Clinical Features
The incidence of chickenpox is not well established since it is not a reportable disease. However, incidence was estimated at between 1.6 and 4.6 per 1,000 individuals between 15 and 44 years of age in the United States during the 1990s [49]. In varicella, there is a generalized rash with lesions that begin as small erythematous macules and papules, and then evolve into vesicles, pustules, and crusted papules (Fig. 12.4). Lesions tend to be more densely distributed on the trunk than the extremities, and can range in number from very few to many lesions.
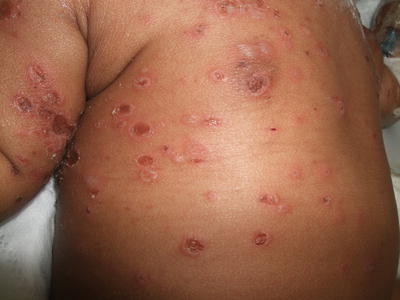
Fig. 12.4
Varicella zoster lesions present as erythematous vesicles and erosions in various stages of development on the trunk
Herpes zoster or shingles is a secondary varicella zoster virus disease caused by reactivation of latent viral infection. Lesions are typically erythematous macules and papules that become vesicular or pustular, and then scab over. The classic eruption is largely unilateral, and tends to occur in a dermatomal distribution .
12.4.2 Histology
The histologic changes seen with varicella zoster are identical to those seen with herpes simplex and herpes zoster infections (See Sects. 12.1 and 12.3) (Figs. 12.5 and 12.6). These entities cannot be differentiated with routine histologic sections.
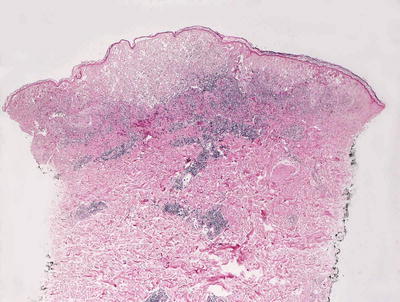
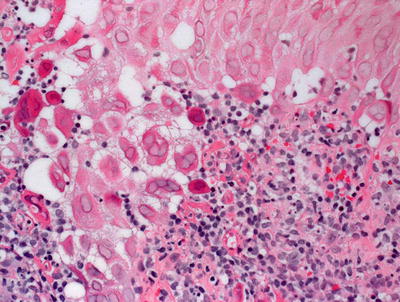
Fig. 12.5
Varicella zoster demonstrates an epidermis that is focally disintegrating secondary to balloon degeneration of keratinocytes
Fig. 12.6
Herpetic viral inclusions are present in varicella zoster . Multinucleated cells, nuclear molding and clearing can be seen in most cases
12.4.3 Pathogenesis
Varicella zoster virus, the causative agent of chickenpox and shingles, is a member of the alpha herpesvirus family of double-stranded DNA viruses. The virus is transmitted by direct contact or airborne infection [57, 58]. T lymphocytes are infected by spread of the virus from the inoculation site at mucosal epithelium. T lymphocytes that are infected with varicella zoster virus undergo a surface remodeling resulting in activation of T-cell receptor and the PI3-kinase pathway [59]. Infected T cells transport viral particles to the skin and other target sites [59]. Nerve ganglia serve as repositories for the virus. In certain circumstances that are not fully understood, varicella zoster virus begins to replicate and migrate down sensory nerves into the skin with concomitant cutaneous vesicular eruption and pain [60]. In children, varicella zoster is most frequently encountered in the setting of immunosuppression [61].
12.5 Congenital Rubella
12.5.1 Clinical Features
Rubella virus infections (German measles) in a healthy child or adult generally cause a mild illness characterized by a diffuse morbilliform eruption. However, rubella infection during the first trimester of pregnancy can cause fetal loss or fetal defects in up to 90 % of cases. There are approximately 100,000 cases of congenital rubella syndrome worldwide each year [62]. Fetal abnormalities are rarely seen with infections that occur after the 16th week of pregnancy, although hearing loss has been reported with infections up to 20 weeks. The organs most commonly affected are the eyes, ears, heart, and brain. Affected children are also at increased risk for developmental delay and disability.
12.5.2 Histology
It is rare to see biopsies from patients with cutaneous manifestations of congenital rubella, and published reports of histologic findings are uncommon. The histologic changes are nonspecific, and consist of a superficial and deep perivascular lymphohistiocytic infiltrate [63]. The infiltrate can be quite dense, extend into the subcutis, and consist of B lymphocytes deep in the infiltrate [64]. Epidermal changes are not present, and no viral inclusions are identified. On electron microscopic examination, abundant tubuloreticular structures are seen within capillary endothelial cells, fibroblasts, and dermal lymphocytes [64].
12.5.3 Pathogenesis
Rubella virus belongs to the Rubivirus genus of the Togaviridae family of single-stranded RNA viruses [62, 65]. Rubella virus occurs by respiratory transmission from person-to-person, or transplacentally [65]. Proteins located on the external surface of the virus facilitate endocytosis [62]. Once host cells are infected, the virus induces cell-cycle arrest by generating tetraploid nuclei [65]. This leads to cell apoptosis, which may be one of the initiating events that result in the fetal birth detects seen with congenital rubella .
12.6 Cytomegalovirus
12.6.1 Clinical Features
In the United States, approximately 33,000 infants per year are born with congenital cytomegalovirus (CMV). However, only about 10 % of these cases are symptomatic at birth [66]. Cutaneous features of congenital CMV infection are variable, and may include petechiae or purpura, a morbilliform rash, or a “blueberry muffin” appearance.
12.6.2 Histology
It is rare to biopsy a skin eruption resulted from direct cutaneous infection by CMV. When this situation occurs, which is most common in the setting of immunosuppression, large cytoplasmic eosinophilic inclusions of CMV are commonly seen in endothelial cells [67, 68]. This leads to vascular thrombosis, vasculopathy, and occasionally vasculitis with hemorrhage and variable amounts of peri-vascular inflammation [69]. Viral inclusions have been described in dermal fibroblasts, histiocytes, eccrine apparatus [67, 73] and dermal nerves [74]. CMV has also been associated with scleroderma-like changes in some patients [68]. In children, CMV has been implicated in the pathogenesis of acute hemorrhagic edema, in which abundant dermal edema accompanies changes of leukocytoclastic vasculitis [70–72].
12.6.3 Pathogenesis
CMV is a member of the beta-herpesviridae subfamily [75]. Cutaneous manifestations of CMV are most frequently encountered in neonates as a result of in utero transmission [76]. The virus infects epithelial cells, hematopoietic cells, endothelial cells and connective tissue, and may have a prolonged latency period [75]. Viral glycoproteins play a key role in specific cellular entry and infection [77]. The virus persists by virtue of its “immune evasion genes,” which allow it to escape from immune cells and neutralizing antibodies [78]. The high level of genomic diversity in CMV enables reinfection by heterotypic strains. Invasion of endothelial cells with subsequent cellular damage, loss of vascular integrity, and thrombosis can be directly attributed to CMV infection in patients with cutaneous manifestations [69].
12.7 Orf
12.7.1 Clinical Features
Orf is a skin infection caused by parapoxvirus that typically infects goats and sheep [79]. Human infections usually occur in teens and adults with occupational exposure, although cases have been reported in children who visit petting zoos or livestock fairs. Lesions usually occur on the hands or fingers, and start as small erythematous papules that enlarge to form tender nodules which can ulcerate and crust over. The natural history of orf is spontaneous resolution over a few weeks .
12.7.2 Histology
Early lesions demonstrate a moderate degree of epidermal acanthosis with significant pallor within keratinocytes at superficial levels of the epidermis (Figs. 12.7, 12.8 and 12.9). Within these cells, viral cytoplasmic inclusions are seen. Intraepidermal vesicles are often present secondary to marked edema within the dermis. The dermis demonstrates a proliferation of thin-walled vessels with a surrounding infiltrate of lymphocytes, histiocytes, and rare eosinophils and plasma cells [80, 81]. As the lesions evolve, epidermal necrosis is almost invariably present, often prominent in the central portions of the lesions. Neutrophils are present adjacent to the necrotic foci. Viral particles are readily apparent on electron microscopy [81].
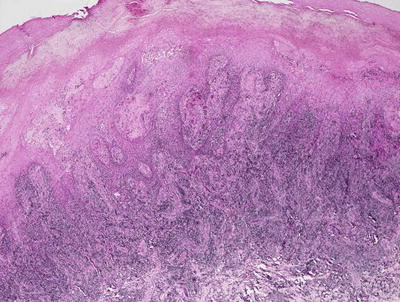
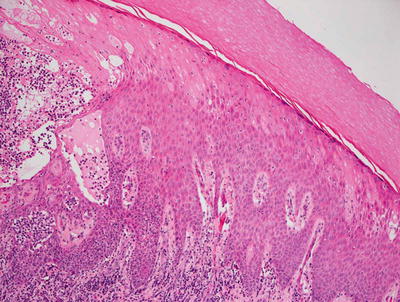
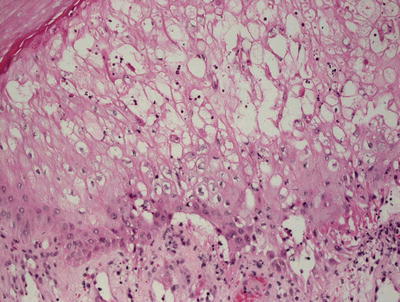
Fig. 12.7
An acanthotic and spongiotic epidermis overlies a dense lymphohistiocytic inflammatory infiltrate in orf
Fig. 12.8
Prominent epidermal spongiosis and dense inflammation are seen in orf
Fig. 12.9
Dissolution of the epidermis and eosinophilic cytoplasmic inclusions resembling erythrocytes are present in orf
The differential diagnosis includes a milker’s nodule that is histologically identical to orf, but can be differentiated by electron microscopy. Early lesions may resemble spongiotic dermatitis, but the epidermal pallor helps to make this distinction. The presence of viral particles establishes the diagnosis. The epidermal pallor may also be seen in nutritional disorders, although the clinical presentation is markedly different, and these entities do not have cytoplasmic inclusions within keratinocytes. Epidermal acanthosis along with proliferation of small well-formed capillaries may raise the diagnostic possibility of a pyogenic granuloma. Again, the presence of epidermal pallor and cytoplasmic viral inclusions are helpful in making the distinction .
12.7.3 Pathogenesis
Orf is a DNA parapoxvirus that is transmitted to humans through direct contact with infected goats and sheep [82]. The virus expresses an anti-apoptotic factor (similar to Bcl-2) and develops interferon resistance that enable it to survive in the host. It also expresses a soluble factor similar to interleukin-10 that suppresses pro-inflammatory cytokines, and a chemokine-binding protein that inhibits immune cell tracking [81, 83].
12.8 Milker’s Nodule
12.8.1 Clinical Features
Milker’s nodules usually affect the hands of individuals who work with cattles. One or aseveral small red papules appear and develop into large and tender nodules. The nodules spontaneously resolve without scarring over a few weeks.
12.8.2 Histology
Histologic features vary with the stage of the lesion biopsied, and they are histologically indistinguishable from the changes seen in orf. The viruses can be visualized by electron microscopic examination.
12.9 Molluscum Contagiosum
12.9.1 Clinical Features
The classic cutaneous lesion of molluscum is a small waxy, skin-colored, dome-shaped papule that may have a central dell or umbilication (Fig. 12.10) [86]. At times, lesions become inflamed with swelling, erythema, purulent discharge, and crusting. Molluscum infection can trigger development of an eczematous dermatitis with erythema and scale surrounding the affected areas.
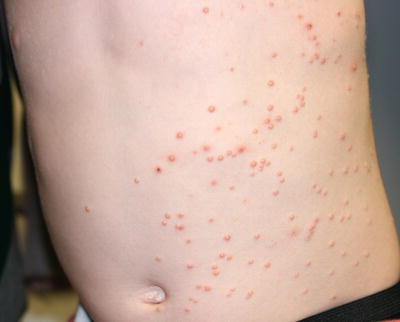
Fig. 12.10
Multiple domed, erythematous and skin-colored, umbilicated papules are seen on the trunk in molluscum contagiosum
12.9.2 Histology
Histologic changes include a lobular proliferation of keratinocytes that contain eosinophilic cytoplasmic inclusions with a very characteristic appearance (Figs. 12.11 and 12.12). The inclusions begin in suprabasilar keratinocytes, and become progressively larger and more basophilic with upward progression in the epidermis. The epidermis is acanthotic with variable amounts of spongiosis. Transepidermal elimination is seen in many cases. An intense inflammatory response is present, and this is accentuated following rupture of the lesions. In some cases, atypical CD8+ lymphocytes and CD30+ cells are present within the infiltrate, leading to concern for lymphoma [10, 87]. Similarly, an intense inflammatory response has been identified in patients with altered immune systems [88]. Follicular involvement is commonly seen in immunosuppressed patients [4, 89].
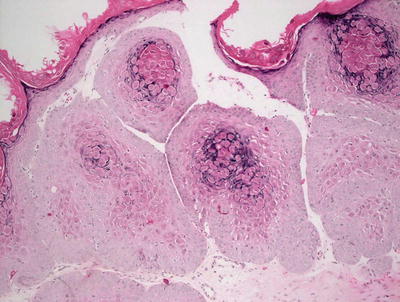
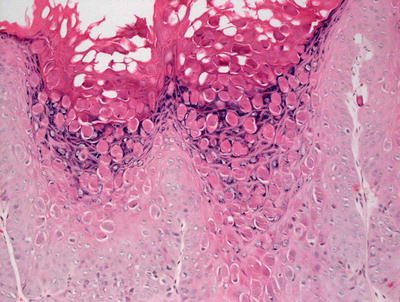
Fig. 12.11
The epidermis is acanthotic and has a lobular appearance with abundant viral inclusions within keratinocytes in molluscum contagiosum
Fig. 12.12
Eosinophilic viral inclusions are present in keratinocytes and extruded at the epidermal surface in molluscum contagiosum
12.9.3 Pathogenesis
Molluscum contagiosum is caused by molluscipoxvirus, which is a double-stranded DNA virus in the family Poxviridae [86]. The virus induces cell mitosis and disrupts epidermal cell differentiation. It is limited to the epidermis, and replicates within the cytoplasm of keratinocytes. Molluscipoxvirus upregulates epidermal growth factor and interferes with cell signaling pathways that activate nuclear factor-κB, thus inhibiting host immune response [90, 91].
12.10 Eczema Herpeticum/Kaposi’s Varicelliform Eruption
12.10.1 Clinical Features
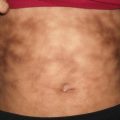
Eczema herpeticum (Kaposi’s varicelliform eruption) is a widespread cutaneous infection with herpes simplex virus occurring in the setting of preexisting chronic skin disease, usually atopic dermatitis [92]. Patients present with many small monomorphous vesicles on an erythematous background. The lesions typically spread rapidly over the skin and evolve into vesiculopustules, crusted papules, and superficial “punched out” erosions (Fig. 12.13). Patients may also have fever, malaise, and lymphadenopathy, and they often require hospitalization for intravenous fluids, medications, and proper skin care.
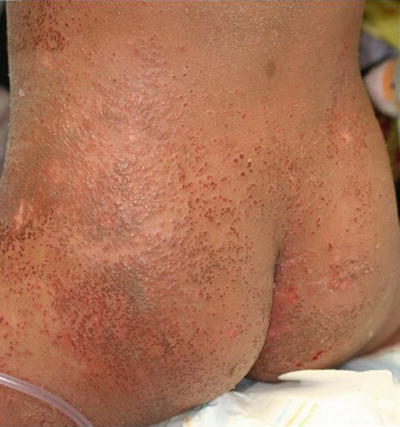
Fig. 12.13
Monomorphic, punched out erosions overlie a generalized eczematous eruption on the lower trunk in eczema herpeticum
12.10.2 Histology
Histologic changes in eczema herpeticum are those of a herpetic dermatitis superimposed upon mild spongiotic dermatitis. Multinucleation, nuclear molding, margination of chromatin, and acantholysis are characteristics of herpes virus infection seen in the epidermis (Figs. 12.14 and 12.15). These changes reflect the nature of the condition that is essentially a generalized infection with herpes virus in the setting of atopic dermatitis. Cowpox and vaccinia infections may demonstrate a similar clinical appearance, but the changes on biopsy vary based upon distinct histologic features specific to the offending viral agent. Similar clinical eruptions have been described in patients with Darier’s disease , Hailey-Hailey disease , rosacea, and mycosis fungoides, the changes are not specific [93–95]. In these cases, histologic evidence of the underlying conditions is apparent when the appropriate clinical sites are biopsied.
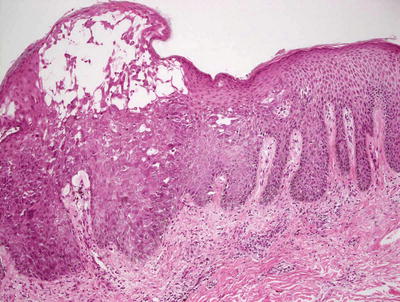
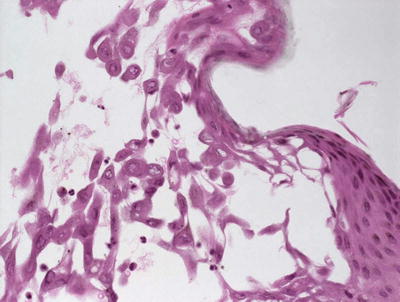
Fig. 12.14
A spongiotic epidermis has blistering with acantholysis and viropathic changes in eczema herpeticum/Kaposi’s varicelliform eruption
Fig. 12.15
Viropathic changes of herpes virus infection, including acantholysis as well as nuclear molding and clearing, are seen in eczema herpeticum/Kaposi’s varicelliform eruption
12.10.3 Pathogenesis
Eczema herpeticum is caused by dissemination of herpes virus or vaccinia virus in the setting of an underlying cutaneous eruption [96]. Cases may also result from infection with coxsackie A16 virus [97]. The viral spread is believed to be due to humoral and cellular immunity dysfunction. Decrease in inflammatory cytokines, such as interferon and CXCL10 as well as altered type 1 T-helper cell immune response, have been observed in affected individuals [98]. Patients also have increased levels of interleukin-4, which inhibits γ-interferon activity [99].
12.11 Verruca Vulgaris
12.11.1 Clinical Features
Common warts (verruca vulgaris) tend to be caused by human papilloma virus (HPV) types 1, 2 and 7. Disease prevalence is estimated at 3.5 % in adults and up to 33 % in school-aged children [100]. Common sites of infection are the fingers and hands, although they may appear in other anatomic sites. Individual lesions appear as hyperkeratotic skin-colored papules and plaques. When present on the face, they are often slender filiform papules.
Plane warts or flat warts are common in children and women, but rare in men. They are most often caused by HPV types 3 and 10. Lesions typically appear on the face, dorsal hands, and shins. Individual lesions are small thin, flat-topped, tan to skin-colored papules that tend to Koebnerize in areas of trauma .
12.11.2 Histology
In verruca vulgaris, focal parakeratosis is present at the tips of exophytic papillomatous proliferations of the epidermis. The epidermis is acanthotic, and demonstrates hypergranulosis with focal clumping of keratohyalin granules (Figs. 12.16 and 12.17). In some cases, there is a slight pallor to the keratinocytes in the mid-portions of the epidermis. At low magnification, slight cup-shaped inward turning of the rete ridges at the periphery of the lesion may be apparent, and these features can be a useful diagnostic clue. There are dilated capillaries within the dermal papillae, and may result in foci of hemorrhage and thrombosis within the stratum corneum.
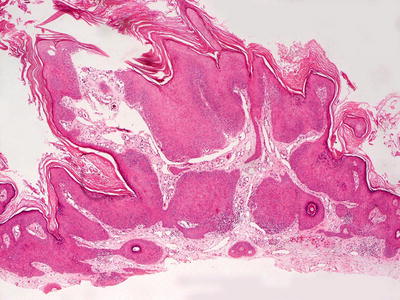
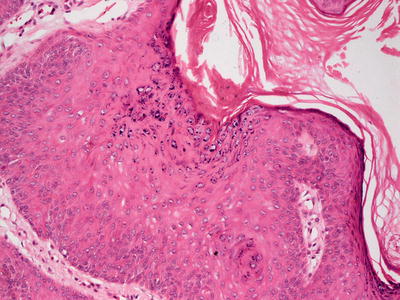
Fig. 12.16
Verruca vulgaris demonstrates an acanthotic and papillomatous epidermis
Fig. 12.17
Clumping of keratohyaline granules within the granular layer is characteristic of verruca vulgaris
The differential diagnosis includes epidermal nevus, seborrheic keratosis, acanthosis nigricans, and trichilemmoma. Epidermal nevi ordinarily do not have parakeratosis at the tips of the papillae, or intervening orthokeratosis as seen in verruca vulgaris. They also do not have inward turning rete ridges, epidermal pallor, and clumping of keratohyaline granules. Seborrheic keratosis and acanthosis nigricans have some similar histologic changes to verruca vulgaris, but they can be easily distinguished based upon clinical history. Trichilemmoma shares many histologic features with verruca vulgaris, but it can be differentiated based upon a characteristic thickened and eosinophilic basement membrane that underlies pale keratinocytes. However, in some cases, it is not possible to make the distinction between these two entities with absolute certainty.
12.11.3 Pathogenesis
Verruca vulgaris is caused by infection with human papillomaviruses (HPV), which are small DNA viruses of the papovavirus family. Infection by HPV requires inoculation of the virus into basal epithelial cells. Active viral DNA replication has been observed to occur in the spinous layer of the epidermis, and the assembly of live virions occurs in the granular layer.
Over 120 HPV subtypes have been identified based on their distinct DNA properties [101]. For practical purposes, HPV are classified as cutaneous and mucosal subtypes according to their tissue tropism. Cutaneous HPV subtypes are those common in the general population (such as HPV-1, 2, and 4) and those found in epidermodysplasia verruciformis (such as HPV-5 and HPV-8 that are high risk for inducing malignant transformation) [100]. They are characterized as low risk, with additional subtypes including types 27 and 57 [102–104]. Mucosal HPV subtypes are classified as low-risk and high-risk types. Low-risk HPV (such as HPV-6, 11, and 53) are usually not found in squamous cell carcinoma. In contrast, high-risk HPV (such as HPV-16, 18, 31, 33, 35, 52, 58, and 67) are often associated with both cutaneous squamous cell carcinoma and cervical cancer [100, 104].
The HPV viral genome encodes for seven to nine open reading frames, depending on the viral subtype. The coding region encodes early and late proteins. Early proteins, including E1–E7, are important in gene transcription, DNA replication, and cellular transformation [100, 105]. E1 and E2 proteins, in combination with various cellular factors, mediate viral replication. Some of the early proteins, such as E4, can disrupt the cytokeratin network in keratinocytes, leading to the morphological viral-induced changes of “koilocytosis” in the affected cells. Some late viral proteins, such as L1 and L2, form viral structures including the viral capsid.
The host immune response is important in the elimination of viral particles. The immune mechanism is partially dependent on cell-mediated immunity with lymphocytes infiltrating viral lesions. The host innate immune response via toll-like receptors (TLR) , interferon-β, and tumor necrosis factor-α are also important in the initial recognition of viral particles by host immune cells and the subsequent immune reaction [106–109].
Malignant transformation induced by high-risk HPV subtypes requires, at least in part, persistent overexpression of the viral oncoproteins E6 and E7. These proteins affect the host cell cycle and apoptosis, leading to unchecked cell proliferation and transformation. Mechanistically, E6 and E7 interfere with the function of the tumor suppressors p53 and Rb to promote dysregulated cell growth [110–112].
12.12 Verruca Plantaris
12.12.1 Clinical Features
Plantar warts are associated with HPV types 1 and 4. They are common in children and typically affect the sole of the foot. Clinically, there is a hyperkeratotic nodule surrounded by a raised rim (Fig. 12.18). “Black dots” (thrombosed capillaries) can be seen over the surface. They are frequently tender with weight bearing or pressure .
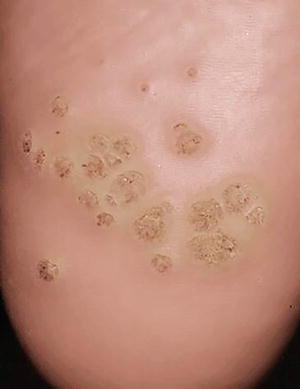
Fig. 12.18
Verruca plantaris presents as skin-colored hyperkeratotic papules and plaques with hemorrhagic dotting on the sole
12.12.2 Histology
Verruca plantaris displays histologic features that can be distinguished easily from verruca vulgaris. The lesions are generally flat. While parakeratosis may be present, it is not a constant finding. The most superficial keratinocytes show vacuolization, and eosinophilic cytoplasmic vacuoles are present in the epidermis, usually sparing the most basilar cells (Figs. 12.19 and 12.20). The cytoplasmic vacuoles are large, and marked keratohyaline granule clumping are readily apparent at low magnification. While there is some epidermal acanthosis, the rete ridges are generally not markedly elongated as seen in verruca vulgaris. In most cases, there is an underlying inflammatory infiltrate, and foci of regression are frequently present in the dermis.
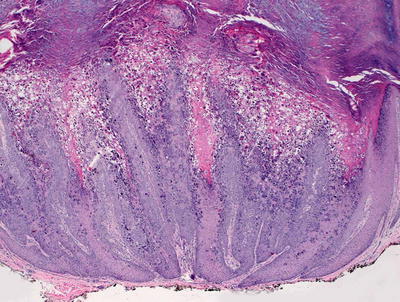
Fig. 12.19
Epidermal hyperplasia , papillomatous growth pattern, and marked keratohyaline granule clumping are seen in verruca plantaris