Fig. 3.1
Plaque psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
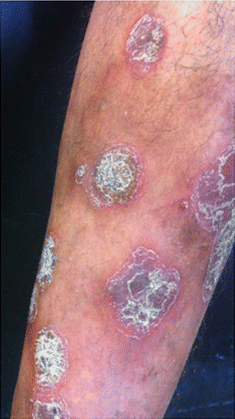
Fig. 3.2
Plaque psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Nail Psoriasis
The cupuliform depressions called pittings of the nails (thimble nails) are the most observed nail changes and occur due to changes of the proximal nail fold. The involvement of the nail plate can determine the appearance of subungual hyperkeratosis, onycholysis and “oil slicks” (brown band between onycholysis and lunula). The longitudinal ridges and onychodystrophy (Fig. 3.3) are caused by damage to ungueal matrix [11].
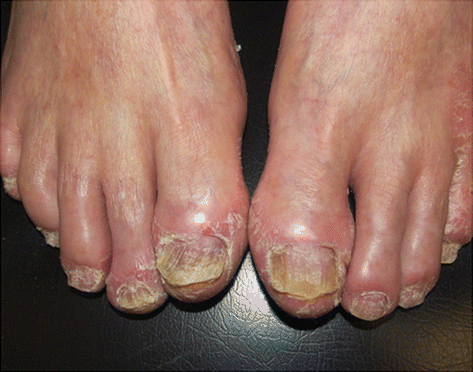
Fig. 3.3
Ungueal psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Guttate Psoriasis
It is characterized by the sudden appearance of erythemato-squamous papules, usually on the trunk and proximal parts of limbs (Fig. 3.4). Being more common in children, teenagers and young adults, this form of psoriasis may be preceded by streptococcic infections and often heals spontaneously in a few months, although in some cases it may persist and evolve to the plaque form [11].
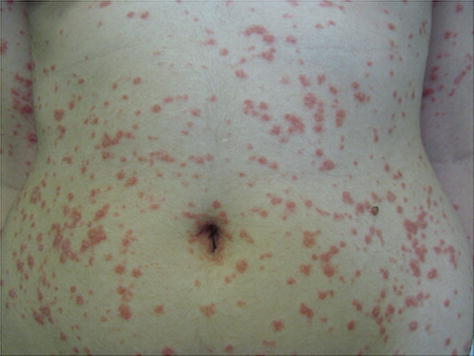
Fig. 3.4
Guttate psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Erythrodermic Psoriasis
An intense erythema is seen over the entire integument, with variable desquamation (Fig. 3.5). Its occurrence may be due to the natural evolution of the disease in immunosuppressed patients (iatrogenic form) or because of the patient’s bad response to treatment. In such a form, it may be systemic and affects the heart, kidneys and liver, in addition to increasing the risk of secondary infections, which requires the patient hospitalization [11].
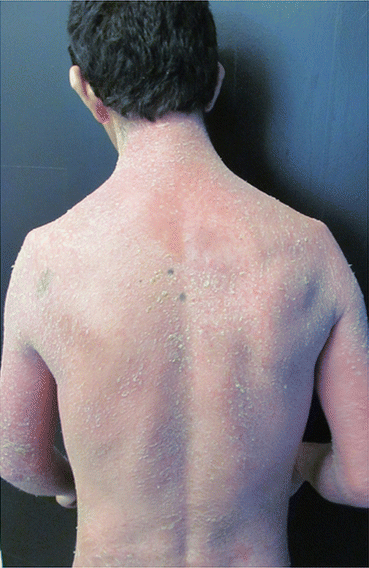
Fig. 3.5
Erythrodermic psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Pustalar Psoriasis
This form of the disease is characterized by the aggravation of both the erythema and edema of previous lesions, followed by the appearance of confluent, non-follicular, superficial pustules, especially on flexures and the trunk (Fig. 3.6). It may be generalized or localized, and evolves from plaque psoriasis because of treatment interruption, infection or hypokalemia, or assumes an idiopathic form. Its occurrence is acute and patients will relapse within a few weeks [11].
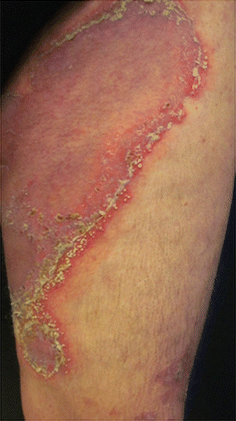
Fig. 3.6
Pustular psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Palmoplantar Psoriasis
This type of psoriasis is more common in adults, being characterized by the appearance of symmetrical, well-defined plaques, showing intense hyperkeratosis. It may form fissures, except on the plantar cavus (Figs. 3.7 and 3.8) [11].
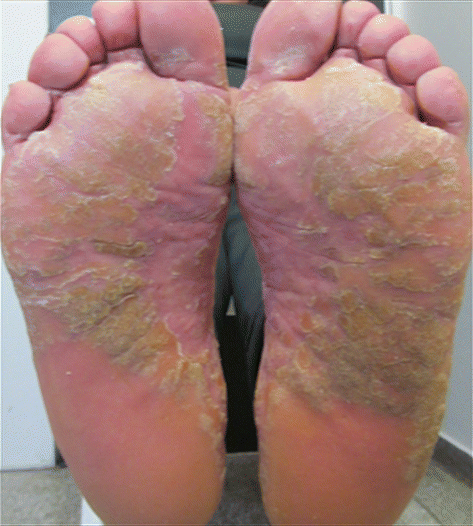
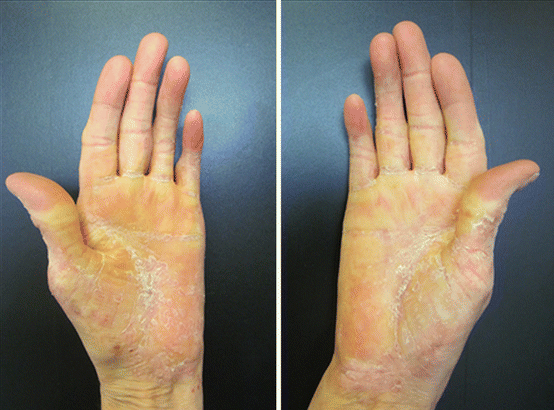
Fig. 3.7
Plantar psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Fig. 3.8
Palmar psoriasis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Psoriasis in Children
Atypical presentations may be observed in infants, but it is mostly characterized by a few erythematous plaques with little desquamation, occurring in the perioral, periorbital, the scalp and genital regions (Fig. 3.9). Follicular affection is common. There is no correlation between an early start of the disease and the worst prognosis [11].
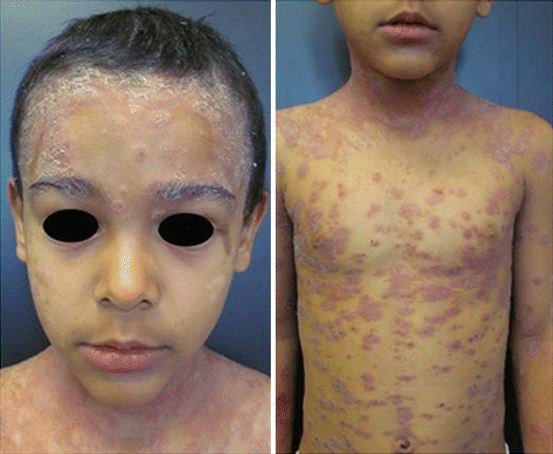
Fig. 3.9
Psoriasis in children. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Psoriasis in the Elderly
Lesions are mainly observed in inferior limbs and the scalp, with pronounced pruritus. In general, the disorder varies from mild to moderate, but treatment may be impaired by drug interactions [11].
Anatomopathology
In psoriasis, histopathological findings are nonspecific but very characteristic, and may be found in other dermatological diseases, such as in Pityriasis Rubra Pilaris and Lichen Simplex Chronicus [12]. In initial lesions, vascular proliferation and superficial vasodilation are observed in the dermis (Fig. 3.10). The perivascular infiltrate is composed of lymphocytes and macrophages. With plaque development, this infiltrate becomes a mixture of lymphocytes, macrophages and neutrophils, with neutrophils and lymphocytes migrating towards the epidermis. Vascular alterations become even more evident, with an increase in the number and size of capillaries, as well as the acquisition of a tortuous appearance (Fig. 3.11) [13]. In the epidermis, there is an increase in the spinous layer (acanthosis), as seen by the regular elongation of the dermal papillae and epidermal cones (Fig. 3.12). The suprapapillary region becomes thinned and the granular layer may be reduced or absent (hypogranulosis or agranulosis) [12]. In the corneal layer, hyperkeratosis with parakeratosis (because of the increase in nucleated keratinocytes) is evident. Neutrophils that migrated towards the epidermis gather forming Munro’s microabscesses in the corneal layer, and spongiform pustules of Kogoj in the spinous layer [12].
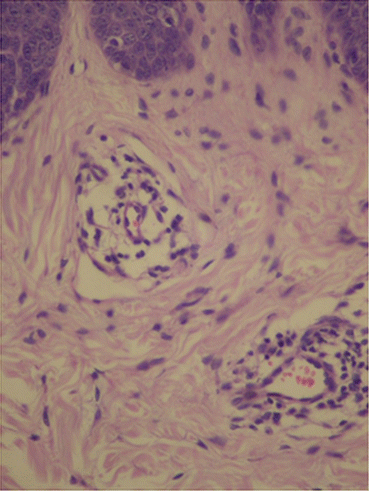
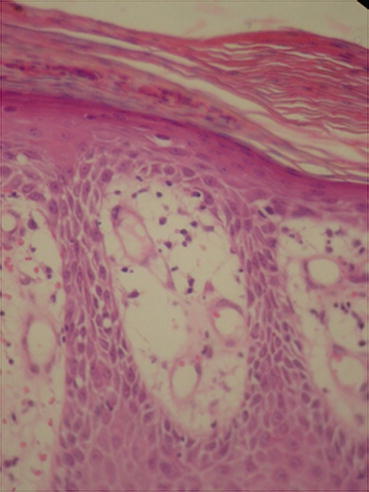
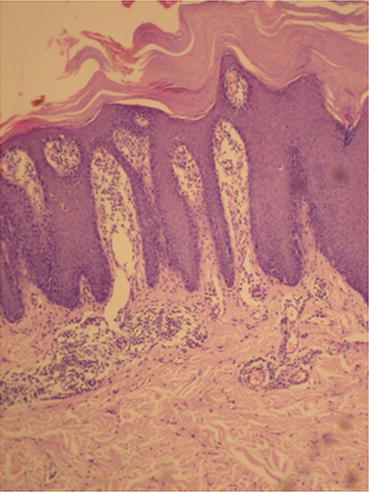
Fig. 3.10
Vascular proliferation and vasodilation in the superficial dermis. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Fig. 3.11
In the superficial dermis, capillaries are increased in number and size, and show a tortuous appearance. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Fig. 3.12
Acanthosis, elongation of dermal papillae, hyperkeratosis with parakeratosis, and thinning of the suprapapillary region. Archives of the Ambulatório de Psoríase, Pontifícia Universidade Católica de Campinas
Angiogenesis in the Etiopathogenesis of Psoriasis
In spite of the advances in treatment for this chronic, immune-mediated, inflammatory disease, there are still etiological aspects of psoriasis that are not completely understood [14].
Specific cells responsible for triggering psoriasis have not been determined yet, although it is known that T-helper lymphocytes (especially T-helper-1 and T-helper-17) and T regulators contribute to the pro-angiogenic environment fundamental to the beginning and maintenance of the disease [14, 15].
Angiogenesis represents one of the initial events in psoriasis, occurring even before plaque formation [13].
Angiogenesis is the process by which new capillaries form from pre-existing vessels, with the use of the extracellular matrix and the recruitment of pericytes and smooth muscles cells. It occurs physiologically during embryogenesis, the feminine menstrual cycle, and in tissue repair and cicatrization processes. Inflammatory diseases, such as psoriasis, and the development of tumors are also marked by angiogenesis.
The papillary dermis of patients with psoriasis presents peculiar histological features, including elongated, tortuous, blood capillaries, increased in diameter and number [16–19]. Such vascular alterations occur prematurely and precede the characteristic epidermal hyperplasia of the disease [17, 20]. Effective treatments normalize this vascular dilation before alterations on the epidermis are seen, but other structural modifications in neo-formed vessels may persist for up to 9 months after clinical improvements are observed [19]. Increases in the cutaneous blood flow caused by angiogenesis are also observed in the clinically healthy skin, surrounding the plaque [21, 22].
Clinically, the dilation and elongation of blood capillaries correspond to the Auspitz’s sign. This sign is characterized by the presence of small bleeding points resulting from the mechanical rupture of the neo-formed vessels in the papillary dermis, after the scales have been removed through Brocq’s methodic curettage [23].
In addition to the structural alterations of vessels seen in the papillary dermis, there is an increase in the migration of inflammatory cells and in the expression of adherence molecules, such as E-selectin, ICAM-1 (Intercellular Adhesion Molecule 1) and the vascular cell adhesion molecule-1 [24, 25].
The strong adhesion of leucocytes to endothelial cells triggers the inflammatory response, coordinated by the infiltrate composed of neutrophils, mastocytes and especially of (CD4+) T-helper lymphocytes, which secrete inflammatory cytokines such as IL-17, tumor necrosis factor (TNF) and gamma interferon (IFN-γ) [7, 26, 27]. In the presence of IFN-γ, the immune response becomes polarized. As a result, a pattern of Th1 and Th17 lymphocytes is formed, an important event in the physiopathology of psoriasis for these cells are responsible for the proliferation of keratinocytes in the epidermis [1, 13].
A study with keratinocytes of both lesioned and non-lesioned skin showed that endothelial cells of sick patients exhibited a greater stimulus to migrate, when compared to those of healthy individuals [28].
During angiogenesis, endothelial cells use the components of the extracellular matrix to migrate [13]. Such a mechanism is mediated by transmembrane, heterodimeric proteins expressed in their walls, the integrins [13]. These proteins are responsible for the activation of intracellular signaling pathways [13] and modulate the pro-angiogenic response [29]. Among integrins, αvβ3 is expressed in low levels in the normal vascular tree, acting as a receptor for the von Willebrand factor, fibrinogen and fibronectin [30]. As a consequence of the inflammatory process, the expression of αvβ3 is highly increased during angiogenesis [31–34]. In psoriasis, the expression of this integrin increases in endothelial cells compared to the skin of healthy individuals [35, 36].
Once angiogenesis is one of the main features of psoriasis, several studies have focused on the identification of pro-angiogenic mediators in the affected skin, which revealed a vast spectrum of factors, among which the vascular endothelial growth factor (VEGF), hypoxia inducible factors (HIFs), angiopoietins, the tumor necrosis factor (TNF), interleukins (IL-8, IL-17) and the transforming growth factor-β and α (TGF-β and TGF-α) [13, 37].
Vascular Endothelial Growth Factors (VEGF)
The development and activity of psoriasis are directly correlated to VEGF levels and their high-affinity receptors VEGFR-1 and VEGFR-2 [13, 14]. The active form of VEGF is a 40-45 kDa, homodimeric glycoprotein, described for the first time as a vascular permeability factor [13, 38].
Twelve subtypes are known, seven of which show pro-angiogenic properties, and five anti-angiogenic action [39]. The gene encoding one of these subtypes, VEGF-A, is located near the PSORS1 gene (associated to the hereditary origin of psoriasis), both on chromosome 6p21 [40]. This gene is highly polymorphic, with some forms being associated to the early development of psoriasis [15]. Among the four isoforms (VEGF-A121, VEGF-A165, VEGF-A189, VEGF-A206), VEGF-A165 is the most common to stimulate angiogenesis [15] (Table 3.1).
Table 3.1
VEGF-A
Isoforms | Functions | Secretion | High-affinity receptorsb |
|---|---|---|---|
VEGF-A121 | 1-increase vascular permeability | Keratinocyte stimulation by the tumor growth factor (TGF)-α and external skin trauma. | VEGFR-1 |
VEGF-A165a | 2-control differentiation of endothelial cells | VEGFR-2 | |
VEGF-A189 | 3-monocyte activation and chemotaxis | ||
VEGF-A206 | 4-keratinocyte proliferation. |
VEGF can increase the expression of their own receptors on both keratinocytes and endothelial cells, which results in the activation of tyrosine kinase-mediated signal transduction pathways [15, 41–43]. One of these receptors, VEGFR-2, was shown to be highly associated to the proliferation, migration and the increased vascular permeability induced by VEGF [44].
The expression and secretion of skin VEGF-A by keratinocytes are triggered by the tumor growth factor (TGF)-α, found in high concentrations in patients with psoriasis [15, 45]. Increases in the expression of VEGF m-RNA, VEGF-A itself and its high-affinity receptors, were demonstrated to occur in the epidermal keratinocytes and endothelial cells in the papillary dermis of sick patients [45].
In patients with psoriasis, the serum of both the affected and unaffected skin shows increased VEGF-A levels. Such an increase is correlated to disease severity and favors the early development of symptoms [46–48]. A study with transgenic mice expressing increased levels of VEGF demonstrated the development of psoriasis-like lesions, both clinically and histologically [49, 50]. These mice also remained healthy when treated with VEGF antagonists, corroborating the role of VEGF in the physiopathology of psoriasis [49, 50].
Case reports suggest that VEGF inhibitors, currently approved for the treatment of some tumors and ophthalmopathies, may improve the condition of patients with psoriasis [50]. Given the multifactor nature of this disorder, it is unlikely that VEGF inhibitors will be effective in all cases [50]. However, experimental data on the use of VEGF inhibitors in rats validate its importance for the treatment of particular cases [50].
When secreted physiologically, VEGF not only causes intense angiogenesis, but also contributes to the proliferation of keratinocytes and to the homeostasis of the epidermal layer, thus supporting skin recovery following acute disturbances [13]. Psoriasis may be also induced as a result of external trauma, a response known as the Koebner phenomenon. In addition, the disturbance of the skin barrier can induce simultaneous increases in VEGF concentrations in the skin [51].
Hypoxia Inducible Factors
The cardiovascular system provides both the oxygen and nutrients to tissues. When oxygen levels are reduced, a metabolic response is generated, with the release of Hypoxia Inducible Factors (HIFs) [13]. A physiological response is then triggered, resulting in angiogenesis. HIFs are composed of three α subunits (HIF-1α, HIF-2α, HIF-3α) and one β subunit (HIF-1β) [1].
HIF-1α is the most studied of them. An increase in its expression was observed under low oxygen concentrations, triggering the transcription of target genes associated to hypoxia [1].
Under physiological oxygen concentrations, α subunits are continually synthesized and expressed in the cytoplasm of lymphocytes. Its activity is controlled by degradation processes regulated by the proteasome, a complex of cytoplasmic and nuclear proteins capable of converting proteins into oligopeptides, with ATP consumption. The degradation occurs through the hydroxylation of the prolyl residues of α subunits by prolyl oxidases, which are only active under normal oxygen levels. The hydroxylated form is recognized by the Von Hippel-Lindau (VHL) tumor suppressor, and then finally enters the ubiquitin-proteasome pathway [52, 53].
Under hypoxia conditions, HIFs degradation is interrupted because of the inactivation of prolyl hydroxylases by oxygen reactive species [2]. As a result, HIF-α subunits are not metabolized, leading to the phenomenon of nuclear translocation, in which angiogenesis-related genes are activated [54, 55].
HIFs have many gene targets, including different angiogenesis-regulating proteins: VEGF, angiopoeitins, VEGFR-1, VEGFR-2, IL-8 and Tie-2 [56–64].
An increased expression of those factors is observed in psoriatic lesions. This is probably due to the increase in the metabolic expenses resulting from epidermal proliferation, which leads to tissue hypoxia [1].
Angiopoietins
Angiopoietins (Ang-1 and Ang-2) are endogenous glycoproteins. Their binding to tyrosine kinase receptors (Tie-2) is crucial to the angiogenesis process [65–70]. Ang-1 and Ang-2 are antagonists.
Angiopoietin-1 induces the phosphorylation of the Tie-2 receptor, promoting the activation of an intracellular signal transduction cascade, which results in vessel remodeling and stabilization during angiogenesis [69] (Table 3.2). In embryogenesis, the formation of immature vessels is induced by VEGF. Here, angiogenesis depends on the interaction between VEGF and Angiopoietin-1 to be completed. In adult tissues, low levels of activated Tie-2 guarantee that the mature vascular endothelium remains at rest [71].
Table 3.2
Angiopoetin’s functions
Function | Action on Tie-2 receptor | Expression sites | |
|---|---|---|---|
Angiopoietin-1 | 1. Vascular remodeling and stabilization. 2. Finishes the angiogenesis process. | Induces phosphorylation. | Fibroblasts and mononuclear cells. |
Angiopoietin-2 | 1. Blocks angiogenic stimulation. 2. Vascular regression. 3. Sensitization of endothelial cells to inflammatory stimulation. 4. Leucocyte chemotaxis and adhesion. | Antagonizes phosphorylation. | Endothelial cells. |
Angiopoietin-2, in turn, antagonizes the phosphorylation of the Tie-2 receptor, promoting vascular destabilization, which will lead to vascular regression in the absence of pro-angiogenic stimulation [13, 70] (Table 3.2).
It has been demonstrated that, during psoriasis, the Angiopoietin-Tie-2 system is activated in the papillary dermis [72, 73]. Endothelial cells express Angiopoietin-2, while Ang-1 is expressed by fibroblasts and mononuclear cells [72]. The reduction in Ang-2 expression, following adequate treatments, attests the importance of this glycoprotein in angiogenesis, and its relation to etiopathology of the disease [72].
In a study with transgenic mice expressing increased levels of the Tie-2 receptor, histological alterations similar to those of psoriasis were observed, such as epidermal hyperplasia, hyperkeratosis, parakeratosis, as well as an increase in dermal vascularization [73]. Suppressing this receptor expression completely reversed psoriasis [73].
Angiopoietin-2 is also capable of sensitizing endothelial cells to several inflammatory signals, facilitating leucocyte adhesion and chemotaxis, and contributing to inflammatory responses during the development of psoriasis [74].
Cytokines
Cytokines are also involved in angiogenesis, promoting the proliferation, elongation, increases in tortuosity and the dilation of capillaries. In addition, they also control the expression of other pro-inflammatory mediators, responsible for the development of psoriasis [13]. TNF, IL-8 and IL-17, pro-angiogenic cytokines acting on the physiopathology of psoriasis, are described as follows.
TNF
TNF-α is crucial to the development of psoriasis, once the use of anti-TNF drugs are highly effective [75]. TNF-α levels are increased in both the serum and lesioned skin of sick patients [76]. In lesioned skins, it is secreted by T-lymphocytes and antigen presenting cells [77]. This transmembrane protein is a precursor of several pro-inflammatory events, such as the activation of endothelial cells, resulting in the increased expression of adherence molecules and chemokines, keratinocyte stimulation for the production of other inflammatory mediators, and the activation of both macrophages and dendritic cells in the dermis [78]. Some of the pro-angiogenic factors induced by TNF-α are IL-8, VEGF and the fibroblast growth factor [79]. The impact of TNF on angiogenesis varies depending on its concentration and time of action, being also influenced by other factors, such as VEGF and the platelet activating factor [78, 80, 81]. The pre-formed TNF is stored in dermal mastocytes, and released following inflammatory stimulation [13]. Thus, TNF contributes to angiogenesis through direct stimulation, as well as inducing other pro-angiogenic factors [13].
IL-8
IL-8 is an 8.4 KDa, non-glycosylated protein, composed of two cysteine residues separated by a third amino acid. Its precursor belongs to the CXC chemokine family, and contains 99 amino acids [82]. It is secreted by monocytes, neutrophils, fibroblasts and endothelial cells, following stimulation by IL-1, TNF-α, IL-6, IFN-γ, lipopolysaccharides, oxygen reactive species and other mediators of cellular stress [13]. There are at least two types of specific receptors for IL-8 expressed by monocytes, neutrophils, fibroblasts, endothelial, mesothelial and tumor cells [13]. IL-8 binding to its receptors produces factors with both local and systemic effects, with inflammatory activity being directly related to their concentrations. This cytokine shows pro-angiogenic action and is highly expressed in several chronic inflammatory diseases, including psoriasis [83–85]. However, IL-8 angiogenic effect is independent of its pro-inflammatory activity, since it can stimulate angiogenesis without inflammation [84, 85]. It promotes chemotaxis and the activation of neutrophils and other inflammatory cells, as well as the migration, proliferation and formation of a tube of endothelial cells in vitro [83, 86–88]. This interleukin also inhibits the apoptosis of endothelial cells, stimulating anti-apoptotic proteins (Bcl-2), while inhibiting pro-apoptotic proteins (Bax) [88]. IL-8 and its m-RNA are found in high concentrations in psoriatic lesions, which will decrease following effective treatments [89]. In psoriasis, not only IL-8 induces angiogenesis, but also stimulates the proliferation of keratinocytes and other cells. In spite of these findings, studies reporting the efficiency of anti-IL-8 monoclonal antibodies as a treatment option are still lacking [13].
IL-17
Th17 lymphocytes secrete IL-17 after being stimulated by other cytokines (TGF-β, IL-6, IL-21 and IL-23) [13, 90]. The IL-17 cytokine family is formed of six members, IF-17 A-F, involved in inflammatory and autoimmune diseases, such as psoriasis [91]. Since the IL-17 cytokine family shares several receptors, different cellular targets are achieved during the inflammatory process [90]. The presence of m-RNAs of certain cytokines (IL-17A, IL-17F and IL-22) produced by Th17 lymphocytes in lesioned skins indicates the role these cells play on the pathogenesis of psoriasis [90]. IL-17A triggers the production of chemokines, growth factors and adhesion molecules by different types of cells, such as fibroblasts, epithelial and endothelial cells [13]. Some important cytokines produced are IL-6, IL-8, IL-1, G-CSF, GM-CSF and ICAM-1. IL-17 has pro-inflammatory potential, once it promotes neutrophil chemotaxis and induces granulopoiesis [13]. It also promotes the expression of TNF and IL-1β by human macrophages [92]. Studies with mice demonstrated IL-17A induces the formation of new vessels [93]. Part of IL-17A effect on angiogenesis could be explained by an increased expression of pro-angiogenic factors, including VEGF [13]. Recent studies demonstrated the efficacy of anti-IL17A treatments on patients with psoriasis, who showed important clinical improvements (83% of patients reached PASI 75 following 12 weeks of treatment) [90].
Related posts:
 Infantile Hemangioma: New Insights on Pathogenesis and Beta Blockers Mechanisms of Action
Infantile Hemangioma: New Insights on Pathogenesis and Beta Blockers Mechanisms of Action
 Angiogenesis: General Concepts
Angiogenesis: General Concepts
 Angiogenesis and Nonmelanoma Skin Cancer
Angiogenesis and Nonmelanoma Skin Cancer
 Chemoprevention and Angiogenesis
Chemoprevention and Angiogenesis
 Angiogenesis and Pathogenesis of Port Wine Stain and Infantile Hemangiomas
Angiogenesis and Pathogenesis of Port Wine Stain and Infantile Hemangiomas
 Potential Role of Angiogenesis and Lymphangiogenesis in Atopic Dermatitis: Evidence from Human Studies and Lessons from an Animal Model of Human Disease
Potential Role of Angiogenesis and Lymphangiogenesis in Atopic Dermatitis: Evidence from Human Studies and Lessons from an Animal Model of Human Disease
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


