Fig. 12.1
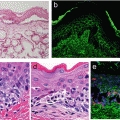
Lymphatic vessels and valves shown by enhanced green fluorescent protein driven by Prox1 promoter in transgenic mice. (a) Cutaneous lymphatic vessels are found in dorsal skin 1 week after birth. Lymphatic vessels are absent in the center median site (arrow) (b) High-power view. Lymphatic vessels show enlargement in the lateral side (arrowheads) and penetration to the median site. (c) Cutaneous lymphatic vessels in the dorsal skin 1 year after birth. Lymphatic vessels are connected at the median site (arrow) (d) High-power image demonstrates the junction of the lymphatic vessels. (e) Lymphatic vessels in mesentery (arrowheads) in transgenic mice. (f) Lymphatic valves are prominently found in mesenteric lymphatic vessels. Scale bars: 1,000 μm (a–e); 500 μm (b–d); 100 μm (f)
12.3 Role of Tumor Lymphangiogenesis in Cancer Metastasis
Lymphangiogenesis is induced in pathological conditions such as tumor progression [12]. Recent studies have identified that several types of cancer including cutaneous malignant melanoma induce new lymphatic vessel growth in the primary site [13, 14]. Note that tumor lymphangiogenesis actively promotes sentinel lymph node metastasis in melanoma, leading to reduced patient survival. VEGF-C, a key growth factor in physiological lymphatic vessel development, was found to induce tumor lymphangiogenesis and enhanced lymph node metastasis in experimental animal models [15, 16]. Tumor lymphangiogensis is also induced in several types of human cancer, and numerous studies found that increased expression levels of VEGF-C are found in the primary sites, and are associated with increased incidence of sentinel lymph node metastasis. The tumor microenvironment recruits several types of inflammatory cells including macrophages, which also express VEGF-C [17, 18].
Tumor-induced lymphangiogenesis was originally identified in experimental animal models. These studies used VEGF-C or VEGF-D as lymphangiogenic growth factor in overexpression models [15, 16, 19]. Moreover, targeted overexpression of VEGF-A in mouse skin also promotes tumor lymphangiogenesis as well as angiogenesis [20]. VEGF-A promotes distant organ metastasis by induction of new blood vessel formation in several types of cancer. Thus, VEGF-A and its receptors are common molecular targets for clinical therapies. Bevacizumab, a neutralizing antibody against human VEGF-A, and several tyrosine kinase inhibitors targeting VEGF receptors are currently utilized for patients undergoing metastatic malignancy such as colon cancer. Thus, lymph node metastasis may be another target for the drugs that functionally impair VEGF receptors and their signal pathways.
12.4 Role of Lymph Node Lymphangiogenesis in Tumor Progression
Lymph node metastasis is associated with reduced overall patient survival in several types of cancer. Primary tumors may induce new lymphatic vessel growth in draining lymph nodes prior to metastasis. Targeted overexpression of either VEGF-A or VEGF-C in mouse skin showed active promotion of lymph node lymphangiogenesis in a standard multistep chemically induced skin carcinogenesis model [20, 21]. It is important to note that lymph node lymphangiogenesis also promoted enhanced metastasis to distant sites, indicative of an advanced tumor progression.
Lymph node lymphangiogenesis represents a premetastatic niche that promotes the initial steps of cancer metastasis. Furthermore, recent studies showed that the premetastatic niche plays a key role in maintaining the survival of cancer stem cells as well as the promotion of metastasis. Lymphatic endothelial cells have been found to suppress immune response in lymph nodes [22]. Therefore, further studies are needed to investigate the significance of lymphatic vessel development in lymph nodes by focusing on the interaction between tumor cells and lymphatic endothelial cells.
12.5 Role of Lymphatic Vessels in Inflammatory Diseases
In contrast to the well-defined role of the blood vascular system in mediating skin inflammation [23, 24], the role of the lymphatic system has been poorly researched and has remained largely unknown. Similar to blood vessels [23], lymphatic vessels have been found to be enlarged in psoriatic skin lesions [25]. However, enlargement of lymphatic vessels might reflect impaired drainage function as well as enhanced fluid uptake, and several diverging functions of lymphatic vessels in inflammation have been proposed. Lymphatic vessels might contribute to inflammatory reactions based on the transfer of antigen-presenting cells to draining lymph nodes, and based on the local release of pro-inflammatory mediators [24]. On the other hand, lymphatic vessels might contribute to the resolution of inflammation by draining inflammatory mediators and cells away from the site of inflammation. Recent studies indicate that lymphatic vessels might indeed mediate anti-inflammatory effects.
The first evidence for this came from a model of chronic skin inflammation , where sensitization of keratin 14 (K14)/VEGF-A transgenic mice with oxazolone, followed by challenge of the ear skin with oxazolone 5 days later, led to a chronic, psoriasis-like skin inflammation [25]. In this model, the lymphatic drainage function was greatly impaired in the chronically inflamed skin [26]. Treatment with an antibody against VEGFR-3 resulted in enhanced inflammatory skin swelling [26]. Conversely, when K14/VEGF-A transgenic mice were crossed with K14/VEGF-C mice, which express the potent lymphangiogenesis factor VEGF-C in the skin and have an increased number of cutaneous lymphatic vessels, the oxazolone-induced inflammatory reaction was reduced and there was no development of the chronic, psoriasis-like skin inflammation [26]. Note that this was associated with improved lymphatic clearance function [26], indicating that enhanced drainage of inflammatory mediators and cells from the inflamed skin contributed to the anti-inflammatory effect. A similar effect was seen when the K14/VEGF-A transgenic mice were crossed with K14/VEGF-D transgenic mice that express the related lymphangiogenic factor VEGF-D in the skin. Intralesional injection of recombinant VEGF-C protein over 7 days also resulted in significantly decreased skin inflammation [26]. These results were supported by the finding that the draining function of lymphatic vessels is reduced in chronic UV damage of mouse skin which includes inflammatory changes [27]. It remains to be investigated whether the observed improvement of diabetic wound healing in mice after VEGF-C treatment [28] might also be related to the anti-inflammatory effects of VEGF-C–induced lymphatic vessel activation in the skin.
Beyond chronic inflammation , activation of lymphatic vessels was also found to reduce acute skin inflammation in mice. Induction of contact hypersensitivity reactions in K14/VEGF-C and K14/VEGF-D transgenic mice, which have an increased number of dermal lymphatic vessels, was associated with reduced tissue swelling [29]. Similarly, the inflammatory response to acute UVB irradiation was also reduced in both genetic mouse models [29]. Intradermal injection of a mutated VEGF-C protein that specifically recognizes VEGFR-3 significantly inhibited the inflammatory cutaneous swelling after a single UV irradiation, associated with reduced numbers of CD11b-positive inflammatory cells [27].
The identification of an important role of lymphatic vessels in limiting inflammation also has importance beyond dermatological diseases. Indeed, it was found that the lymphatic drainage function is impaired in rheumatoid arthritis, and that adeno-associated virus (AAV)-mediated delivery of VEGF-C attenuates joint damage in chronic experimental inflammatory arthritis [30]. Lymphatic vessels are also enlarged in inflammatory bowel disease, and inhibition of lymphatic function with an anti-VEGFR-3 antibody resulted in increased inflammation in an experimental inflammatory bowel disease model [31]. Conversely, very recent results indicate that VEGF-C reduces inflammation in experimental inflammatory bowel disease [32].
In summary, there is increasing evidence for an anti-inflammatory activity of cutaneous lymphatic vessels that show reduced activity in chronic skin inflammation. Therefore, activation of lymphatic function via VEGF-C or other mediators might represent a new therapeutic strategy to treat inflammatory skin diseases [33].
12.6 Perspective
Pathological lymphangiogenesis occurs in tumor progression and chronic inflammation. Several growth factors such as VEGF-A promote new lymphatic vessel growth and alter the physiological function in experimental models. Note that VEGF-C potently improves the lymphatic vessel function in pathological settings. Thus far, the molecular regulation of lymphatic vessel development and the functional regulation by key mediators has been studied in detail. Future investigations are needed to investigate the importance of lymphatic vessel interaction with immune cells during tumor development and inflammation, potentially leading to new therapeutic approaches for the treatment of skin diseases.
References
1.
Tammela T, Alitalo K (2010) Lymphangiogenesis: molecular mechanisms and future promise. Cell 140:460–476CrossRefPubMed
Related posts:
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


