4 Hair Follicle Physiology and Mechanisms of Hair Disorders
Summary
Keywords: hair cycle hair follicle anagen telogen catagen telogen effluvium trichorrhexis invaginata hypotrichosis simplex disease male pattern baldness alopecia areata
Key Points
•The hair follicle cycles through three distinct phases: anagen (growth phase), catagen (regression phase), and telogen (resting phase).
•Hair disorders involve various changes to hair fiber caliber, density, and anagen/telogen duration in the hair growth cycle.
•Hair disorders result from alterations in normal hair biology and can be classified into three main categories: alopecia, hirsutism, and hair shaft disorders.
4.1 Introduction
The hair follicle (HF) is a dynamic miniorgan formed with neuroectodermal–mesodermal interaction and holds significant cosmetic importance in humans. It supports important biological functions such as thermal regulation and protection against injury. HFs also increase the perception of tactile stimuli on the skin, and fulfill important sexual and social communication roles.1,2
Hair is found over most body surfaces except the palms and soles, buccal surface of the lip, and parts of the external genitalia. In addition, hair is variably distributed in different parts of the body. Hair growth is reduced with tiny, almost colorless, hair on most of the body surface (vellus hair), whereas hair is longer and more pigmented on the scalp (terminal hair). Differences also relate to the hair’s form (straight or wavy) and color, which depends on the balance of different melanin types.
In this chapter, we will review the basic physiology of the HF, and subsequently explore some causal mechanisms for the disordered HF using selected examples of hair disorders.
4.2 Overview of Hair Follicle Formation
HF morphogenesis starts at an early embryonic stage. Its proper development and regular cycles involve a strong interplay between Wnt, Notch, Hedgehog, and bone morphogenetic protein (BMP) signaling pathways.3,4
The initiation of HF morphogenesis is an ordered series of mesenchymal–epithelial interactions, with the stages of morphogenesis broadly classified into: induction, organogenesis, and cytodifferentiation (Fig. 4.1). During induction, Wnt-mediated signal transduction first arises in mesenchymal cells directing the thickening of overlying epithelial cells to form a placode. Subsequently, in organogenesis, stabilized placodes signal to underlying dermal cells to proliferate and form clusters of dermal papilla (DP) precursor cells. These dermal condensates then signal back to the epithelial compartment to proliferate and grow downward into the dermis. In cytodifferentiation, follicular epithelial cells envelop the dermal condensate to form distinct DP, which instruct the ectoderm to shape the entire HF through the action of growth factors and morphogens.3,4
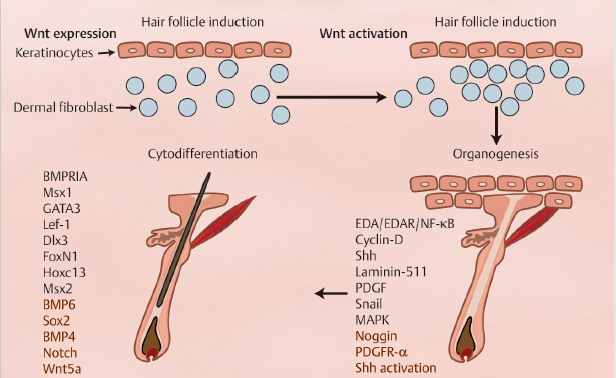
Fig. 4.1 The stages of morphogenesis are broadly classified into: induction, organogenesis, and cytodifferentiation, and its proper development involves a strong interplay between Wnt, Notch, Hedgehog, and bone morphogenetic protein signaling pathways.
Proper positioning and spacing of HFs, on the other hand, is mediated via ectodysplasin receptor Edar-BMP signaling and transcriptional interactions. Edar-directed stabilization of β-catenin is important in determining HF position.
4.3 Hair Follicle Biology and the Hair Cycle
The HF is part of the pilosebaceous unit that also contains the sebaceous gland, apocrine gland, and arrector pili muscle (Fig. 4.2). It is composed of two main compartments: the upper part (infundibulum and isthmus) and lower part (bulb, matrix, and DP).
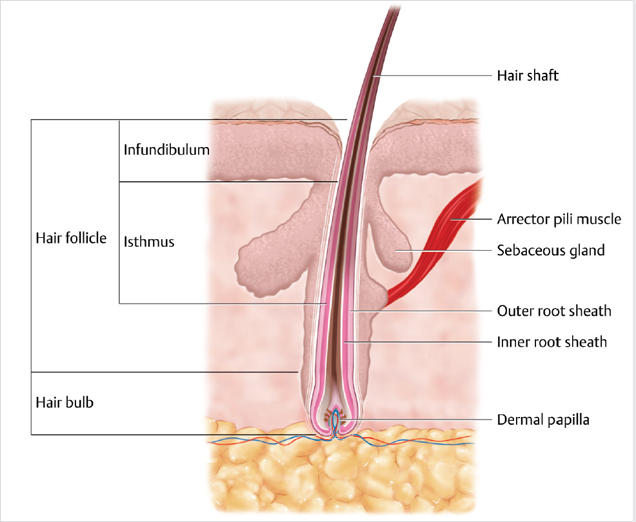
Fig. 4.2 The hair follicle is part of the pilosebaceous unit that contains the sebaceous gland and arrector pili muscle. It is composed of two main compartments: the upper part (infundibulum and isthmus) and lower part (bulb, matrix, and dermal papilla).
The sebaceous gland lies adjacent to the lower part of the infundibulum, and the isthmus contains the hair bulge, which contains the epithelial stem cell reservoir regenerating the HF. Cells migrate from bulge to bulb, where they proliferate and differentiate to produce the hair shaft and all the epithelial cells of the HF. Finally, the inner and outer root sheaths are composed mainly of keratinocytes surrounding the hair shaft.1
The development of hair is dynamic and cyclic, with hormones and cytokines coordinating the duration of growth cycles. This is influenced by factors such as location, age, development stage, and nutritional habits.
HFs grow in repeated cycles, with rapid growth and hair shaft formation alternating with apoptosis-driven HF regression. The characteristic growth and retraction phases that a HF cycles through is known as the hair cycle (Fig. 4.3). This includes three distinct phases: anagen (growth phase), catagen (regression phase), and telogen (resting phase). Exogen (release of the telogen club) does not occur at every cycle. The distinct phases of this process are each regulated by different signals.
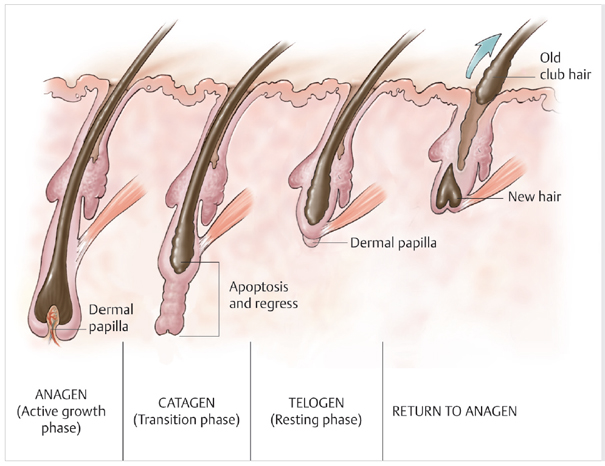
Fig. 4.3 The characteristic growth and retraction phases that a hair follicle cycles through is known as the hair cycle. This includes three distinct phases, namely anagen (growth phase), catagen (regression phase), and telogen (resting phase). (Reproduced with permission from Barrera A, Uebel CO. Hair Transplantation: The Art of Follicular Unit Micrografting and Minigrafting. 2nd ed. Thieme Publishers; 2013.)
4.3.1 Anagen
Anagen is the growth phase, and bulge region stem cells differentiate to all hair lineages, resulting in hair elongation. During this active growth phase, a hair fiber is produced, as the HF enlarges and reaches its characteristic onion shape.
Anagen can be divided into six stages (I–VI). During anagen I–V (proanagen), hair progenitor cells proliferate, envelope the growing DP, grow downward into skin, and begin to differentiate into the hair shaft and inner root sheath (IRS). The newly formed hair shaft then develops, and hair matrix melanocytes show pigment-producing activity. In anagen VI (metanagen), full restoration of the hair fiber–producing unit occurs, and is characterized by formation of the epithelial hair bulb surrounding the DP, which is located deep in subcutaneous tissue, and the new hair shaft appears from the skin surface. The duration of anagen varies according to anatomic location, with this phase in the scalp lasting an average of 3 to 4 years but in some individuals it may last up to 8 years; in contrast, anagen lasts only 3 months in the eyebrow.1,2,5,6
4.3.2 Catagen
Catagen is the regression phase, and starts when anagen ends. During catagen, differentiation and proliferation of hair matrix keratinocytes decreases, melanocytes stop pigment production, and hair shaft production is completed.
At this stage, there is substantial decrease in cell cycling because of increased apoptosis in epithelial cells of the bulb, outer root sheath (ORS), and outermost epithelial layer. Club hairs are formed, with the keratinized brush-like structure at its base anchoring it to the telogen follicle. The DP is transformed into a cluster of quiescent cells closely adjacent to the regressing HF epithelium and travels from the subcutis to the dermis/subcutis border to maintain contact with the distal part of the HF epithelium, including the secondary hair germ and bulge. This phase lasts a few weeks. As majority of HF cells undergo apoptosis, there is shortening of the lower compartment, and DP cells are brought closer to the bulge. This upward movement of the follicular papilla during catagen is crucial for re-establishing follicular papilla–bulge cell contact and induction of a new hair cycle.
Molecules that promote catagen induction have been identified as p75, p53, TGF-β1, FGF5, BDNF, and BMPRIa. It is the exchange of signals between the papilla and bulge that regulates catagen duration. The cells that escape apoptosis during this phase comprise the reservoir that leads to the next anagen.1,2,5,6
4.3.3 Telogen
Finally, telogen begins and the hair goes into resting phase. This phase in the scalp generally lasts 3 months, but may last up to 8 months in some; in contrast, telogen lasts only a few weeks in the eyelashes. Cells enter a quiescent state waiting for signals to re-enter anagen, with an estimated 5 to 15% of scalp HFs remaining in telogen at any time point. Telogen HFs lack pigment-producing melanocytes and IRS. Their DP is closely attached to a small cap of secondary hair germ keratinocytes containing HF stem cells.
At the end of telogen, the hair sheds (exogen). The HF subsequently re-enters the growth phase a few weeks later by stimulating bulge stem cells. The bulge activation theory proposes that bulge stem cells proliferate after signals from the DP. Bulge cell proliferation is the cellular source of the entire HF structure, including hair matrix cells. These daughter cells are transient amplifying cells, which can undergo only a limited number of mitoses, thus establishing the length of anagen and onset of catagen.1,6
Telogen-to-anagen transition is, however, dependent on many factors, and since the HF strongly expresses estrogen receptors during the telogen phase, binding of 17-β-estradiol to these receptors prevents HFs from exiting the telogen phase to enter anagen phase.
Disruption of the processes involved in the hair cycle can therefore lead to various hair growth disorders, and further understanding is essential for the development of more effective therapeutics.
4.4 Hormones and Hair Follicles
The effects of neurohormones on HF growth are complex and strongly dependent on hair cycle stage. A close localization of autonomic and sensory nerve fibers and the bulge area suggests that neuropeptides may influence stem cells and modulate the hair cycle. It is also now clear that HFs are not only a target of neuromediators, but its keratinocytes, melanocytes, and fibroblasts also synthesize neurohormones.
Several studies showed the expression of a neuroendocrine system in the human HF. In particular, the expression of urocortin, corticotropin-releasing hormone (CRH) and CRH receptors, proopiomelanocortin-derived neuropeptides (alpha-melanocyte-stimulating hormone [α-MSH], β-endorphin, adrenocorticotropic hormone [ACTH], thyrotropin-releasing hormone, melatonin), and their associated receptors has been reported.
The role of neurohormones and neuropeptides in human HF pigmentation extends beyond the control of melanin synthesis by α-MSH and ACTH, and includes melanoblast differentiation, reactive oxygen species scavenging, and HF pigmentary unit remodeling.6
4.5 Disorders Associated with Hair Follicles
Hair disorders involve various changes to hair fiber caliber, density, and anagen/telogen duration in the hair growth cycle. These can in turn be classified into three main categories: alopecia, hirsutism, and hair shaft disorders. Alopecia, hirsutism (excessive hair growth in adult women), and hair shaft disorders are caused not only by the malfunction of one or more signaling pathways, but also by other factors such as autoimmune disorders, genetic predisposition, and hormones.3 The following examples of hair disorders highlight various causal mechanisms for the disordered HF.
4.6 Alopecia
4.6.1 Hypotrichosis Simplex Disease
Various forms of alopecia involve deviations from normal signaling pathways. An autosomal disorder, hypotrichosis simplex disease is characterized by sparse/absent scalp hair without structural defects. Affected individuals show normal hair at birth but experience progressive hair loss and thinning of the hair shaft that begins in early childhood. Adenomatosis polyposis down regulated 1 (APCDD1) gene encodes a membrane-bound glycoprotein that acts as a potent inhibitor of the Wnt signaling pathway by interacting with Wnt3A and LRP5. This inhibitory function plays an important role during development of neurons from progenitors; however, the Leu9Arg mutation in the APCDD1 gene inhibits membrane localization and stability, rendering it free to disrupt Wnt signaling pathway, predisposing the patient to alopecia.7
4.6.2 Male Pattern Baldness
Perhaps, the most well-known disorder associated with HFs is male pattern baldness. The main cause of androgenic male pattern baldness is dihydrotestosterone (DHT), which is produced by balding DP. Presence of DHT in HF occurs either through DP capillaries, as it is normally circulated in blood, or by testosterone conversion in balding DP cells. Presence of DHT causes IL-6 upregulation, which promotes expression of IL-6 receptor along with glycoprotein 130 in keratinocytes and matrix cells. The IL-6-upregulated expression results in inhibition of hair shaft elongation and suppression of matrix cell proliferation. IL-6 expression thus induces anagen–catagen progression.8
Another key factor in androgenic male pattern baldness is Dickkopf1 (DKK-1), which promotes catagen. DKK-1 blocks Wnt signaling by preventing β-catenin activation, and activates proapoptotic protein Bax, inducing ORS keratinocyte apoptosis.9 Current research supports that DKK-1 causes anagen to catagen induction, and may play a role in male pattern hair loss.
A few microRNAs have also been implicated, with upregulation of miR-221, miR-125b, miR-106a, and miR-410 in balding papilla cells demonstrated. miRNAs have an established role in the functioning of androgenic receptors, and androgen secretion affects hair color and size via the hair growth cycle. To date, no specific role has been assigned to these miRNAs, and further research is necessary to evaluate their upregulation in male pattern baldness.10
4.6.3 Alopecia Areata
Alopecia areata (AA) is classified as an autoimmune disease of the HF, and recent studies have greatly improved our understanding of its pathogenesis (Fig. 4.4).
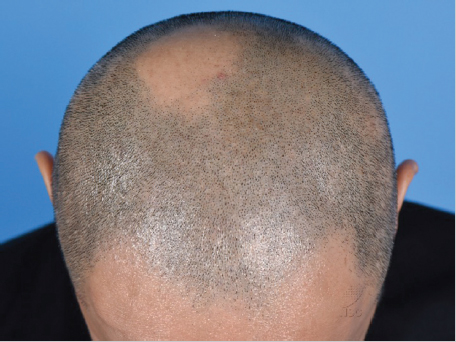
Fig. 4.4 Alopecia areata is an autoimmune disorder of the hair follicle which causes nonscarring alopecic patches over the scalp and other hair-bearing areas of the body.
Related posts:



Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


