Fig. 7.1
Cancer transformation is a multi-stage process and chemopreventive strategies should aim to target the exposure (1), and/or initial cancer formation (2) or (3) cancer promotion
This ROS induced irreversible DNA damage can induce vascular changes, through recruitment of cytokines and chemokines such as vascular endothelial growth factor (VEGF), basic fibroblast growth factor (b-FGF) and interleukin-8 (IL-8) [15]. These discernable and internal changes can further upregulate oncogenic and inflammatory pathways associated with skin cancer including MAPK, EGFR, Src, and p53 mutations [16]. In addition, it can further affect skin appearance, causing erythema, epidermal hyperplasia, vascular dilation and hyperpermeability [17].
Aside from excessive sunlight, people are continuously exposed exogenously whether through food, air, or water to chemical carcinogens and mutagens. Chemical carcinogens are hypothesized to contribute to a significant amount of human cancers, particularly through certain foods and tobacco products. Aflatoxin is one such carcinogen found in the diets of Asian and African communities. Aflatoxin is a mold-produced mycotoxin that directly affects the liver and synergizes with hepatitis B virus. Another chemical carcinogen are heterocyclic amines, produced when muscle meats are heated above 180oC for extended time periods. Though of all the ubiquitously present environmental carcinogens, tobacco products are probably the most notorious. Tobacco products contain a host of chemical carcinogens such as polycyclic hydrocarbons, nitrosamines, and aromatic amines which cause the formation of DNA adducts that lead to miscoding and permanent mutations [18].
Because of its high incidence, skin cancer is one of the most pressing issues in cancer today, and has sparked a need to delay the occurrence of cancer in high-risk populations through dietary or chemical interventions [19]. High-risk populations would include solid organ transplant patients on immunosuppressive drugs, patients with DNA repair defects, as well as those with extensive sun exposure, fair skin, and red hair [20, 21]. Patients who are heterozygous for p16ink4a have multiple atypical nevi and a high incidence of melanoma [22]. Cancers such as melanoma have limited treatment options at late stages, are highly metastatic, and develop rapid drug resistance [13].As a result, scientists conduct research on chemoprevention, the use of synthetic or natural agents that inhibit cellular mechanisms involved in tumorigenesis or before invasion and metastasis occur through prevention, arrest, or reversal [23, 24]. Because cancer transformation is a multistage process, three types of chemopreventive strategies exist: those that inhibit initial cancer formation, those that block cancer promotion, and those that prevent recurrence after anti-cancer therapy [25, 26]. Ideal chemopreventive agents must have little toxicity in healthy populations, and distinctly affect premalignant or malignant cells, rendering normal cells unaffected [27]. In order to develop such drugs researchers must know associated pathways, enzymes, genetic alterations, and cellular targets that are critical to cancer development [28].
Of the tumorigenic pathways researched for chemoprevention, angiogenesis, a hallmark of cancer, is one such targeted event. Angiogenesis is the growth of blood vessels with functions in both normal tissues and cancer cells [16]. In normal tissue, angiogenesis is tightly regulated and fundamental to reproduction, development, and tissue repair [29]. In healthy tissues, vasculature is smaller and abundant with evenly spaced, well-differentiated arteries, arterioles, capillaries, venules, and veins [30]. Tumor vasculature, however, is characterized by a large size with some smooth muscle coating, lack of organization, irregular branching, inconsistent blood flow path and leakiness [28, 30]. These characteristics are a result of hypoxia, inflammation, and oncogenic mutations which make healthy tissues undergo an “angiogenic switch” and upregulate pro-angiogenic agents [31]. For a cancer to grow beyond several millimeters and adopt a metastatic phenotype, tumors must have an autonomous blood supply through neovascularization [15]. This abnormal angiogenesis can lead to zones of ischemia and necrosis as the tumor grows and requires more oxygen and nutrients from the blood supply [30].
The balance between angiogenic factors and angiogenic inhibitors can be disrupted by the aforementioned “angiogenic switch.” These occurrences can cause the increased secretion of angiogenic factors such as cyclooxygenase-2 (COX-2), a pro-inflammatory enzyme, hypoxia induced factor alpha (HIF-1a) a transcription factor that facilitates response to hypoxia, angiopoetin-1 (ANGPT1), an endothelial cell specific growth factor, angiopoetin-2 (ANGPT2), a vascular endothelium remodeler, matrix metalloproteinases (MMPs), extra-cellular matrix remodelers, and basic/acidic fibroblast growth factor (b-FGF or a-FGF), a growth factor involved in cell mitosis and migration, and local immunologic tolerance among others [16, 30, 32, 33]. BFGF is found to be highly expressed in basal cell carcinoma [34] and mutations in FGFR3 are present in some seborrheic keratosis [35]. Though of the numerous promoters of angiogenesis, vascular endothelial growth factor (VEGF) is perhaps the biggest driver. VEGF is an endothelial cell mitogen that when expressed causes blood vessel growth in both normal and abnormal tissue [36]. Aside from cancer, overexpression of VEGF and persistent angiogenesis supports the progression of certain other inflammatory diseases such as ocular neovascularization, which causes blindness, infantile hemangiomas, arthritis, and psoriasis [37–40].
Though the overexpression of a multitude of angiogenic factors is essential for tumor angiogenesis, the suppression of endogenous angiogenic inhibitors is also necessary for tumorigenesis and metastasis. Such endogenous inhibitors include: angiostatin, involved in cell cycle arrest and apoptosis; endostatin, an inhibitor of cell proliferation and migration; canstatin and tumstatin, inducers of apoptosis; arrestin, an inhibitor of VEGF driven angiogenesis and thromospondin-1 (TSP-1) [15, 29, 33].
NSAIDs and COX-2 Inhibitors
Cyclooxygenase (COX), is the rate-limiting enzyme in the metabolism of arachidonic acid to prostaglandins, and exists in two isoforms: COX-1 and COX-2 [41, 42]. COX-1 is constitutively expressed in bodily tissue while COX-2 is induced by certain pathological processes involving inflammation, mitogens, and growth factors [43]. Cyclooxengenase-2 (COX-2) is a vital enzyme that mediates many inflammatory processes, and whose up-regulation is associated with numerous inflammatory disorders and cancers [44]. Animal models have demonstrated the ability for COX-2 to catalyze malignant transformation [45], and is implicated in human cancers with inflammatory origins such as colorectal, gastric, ovarian, cervical, esophageal, and Non-Melanoma Skin Cancer (NMSC) [46, 47]. Tumor promoters, pro-inflammatory cytokines, lipopolysaccharide (LPS), and growth factors can regulate the expression of COX-2 at the transcriptional and posttranscriptional level. While COX-2 is an essential enzyme in the body, its constitutive expression can initiate and promote carcinogenesis [48]. Some cancer-causing substances that initiate this constitutive expression are chemicals in tobacco smoke such as nicotine, nitrosamines, and poly-cyclic aromatic hydrocarbons [49, 50]. These initiators can also come from the environment, the body, and infectious agents such as some fatty acids [51], radiation [52], ultraviolet B [53], free radicals [54], oncogenic proteins [55], growth factors [56], helicobacter pylori [57], HPV [58], hepatitis viruses [59] and Epstein Barr virus [60]. This continuous over-expression of COX-2 promotes carcinogenesis by mutagenesis through increasing reactive oxygen species, mitogenesis through increased production of prostaglandins, angiogenesis by stimulation of vascular endothelial growth factor (VEGF) and platelet-derived growth factor (PDGF), metastasis by increasing production of matrix metalloproteinases (MMP), anti-apoptosis by decreasing arachidonic acid and inhibition of BAX, immunosuppression through the inhibition of B and T lymphocyte proliferation [61] and heightened intracellular telomerase [62].
Since the 1960’s, epidemiological, clinical, and experimental studies have shown the anti-cancer effects of nonsteroidal anti-inflammatory drugs (NSAIDs) through inhibition of COX (Fig. 7.2) [63]. NSAIDs are a family of drugs that are frequently used for inflammation, pain, and fever by reducing the production of specific prostaglandins that promote the aforementioned symptoms along with carcinogenesis [64, 65]. NSAIDs are classified depending on their selective inhibition on the COX isoforms: Selective COX-1 inhibitors, non-selective COX inhibitors, and preferential COX-2 inhibitors [66]. Common NSAIDs like aspirin, ibuprofen, sulindac, and indomethacin inhibit both COX-1 and COX-2 [67] while the later developed coxibs (celecoxib and rofecoxib) selectively inhibit COX-2 [48].
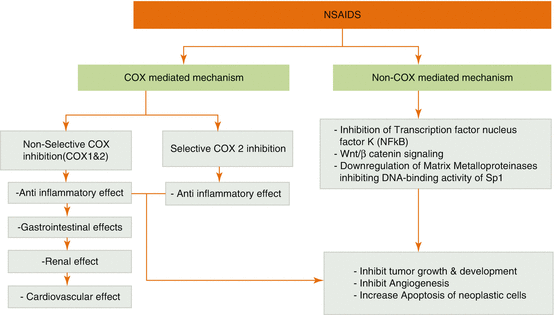
Fig. 7.2
Proposed chemopreventive mechanism of NSAIDS
Preclinical studies have shown that NSAIDs such as sulindac sulfide [68], exisulind [69], and celecoxib [70] can inhibit angiogenesis and cell invasion by inhibiting the DNA-binding activity of Sp1, a driver of VEGF overexpression, and by downregulating MMPs [71, 72]. Studies have also shown that Sulindac can stimulate apoptosis in the normal rectal mucosa of patients with familial adenomatous polyposis coli (FAP) [73] by inhibiting Wnt signaling [74] and reducing β-catenin levels [75]. In addition, sulindac, sulindac sulfide, exisulind, and aspirin can inhibit NF- κB signaling [76, 77]. These agents may also impact the development of NMSC, further proving its efficacy as cancer chemopreventive agents [28].
While COX may be the primary target for NSAIDs, evidently their effect on signaling pathways may also contribute to its chemopreventive properties [24]. Numerous clinical studies have shown the protective effects that NSAIDs have against cancer: Regular intake of 325mg aspirin or 200mg ibuprofen showed a risk reduction of 43% for colon cancer, 25% for breast cancer, 28% for lung cancer, and 27% for prostate cancer [23]. Furthermore, daily intake of selective COX-2 inhibitors, 200mg celecoxib or 25mg rofecoxib, demonstrated a 70% risk reduction in the aforementioned cancers compared to 60% for ibuprofen (COX-1 and COX-2 inhibitor) and 54% for aspirin (COX-1 inhibitor) [23].
However, despite its favorable results, the use of non-selective NSAIDs as a chemopreventive agent is not routinely recommended because of the gastrointestinal and renal side effects that result from the suppression of prostaglandins due to COX inhibition [24]. Therefore, selective inhibition of COX-2 to treat neoplastic proliferation is preferable to non-selective inhibition [78]. By the end of the 90s, the coxib family of drugs was produced to overcome these harsh side effects [79]. Unfortunately, these new drugs caused an increased incidence of cardiovascular side effects after prolonged use [80]. As a result, though clinical studies have shown promising results for using NSAIDs as cancer chemoprevention, long-term administration of COX-2 inhibitors would be ineffective as the risks outweigh the benefits.
Chemoprevention of Basal Cell Nevus Syndrome (BCNS)
BCNS is a rare autosomal dominant disorder characterized by the development of innumerable basal cell carcinomas as well as an increased incidence of medulloblastoma. In addition, other abnormalities are present, including bifid ribs, keratogenic odontocysts, and palmar pits [81].
The major cause of morbidity and mortality in these patients is basal cell carcinoma, which can be locally invasive and rarely metastatic. BCNS is caused by germ line mutations in PTCH, a receptor for the secreted protein Sonic hedgehog. Mutations in PTCH lead to chronic activation of Smoothened and cell proliferation [82]. Smoothened is a transmembrane protein and activator in the Sonic hedgehog pathway [82]. Recently, Smoothened inhibitors have been introduced into the clinic, especially for large and inoperable basal cell carcinomas [83].
These inhibitors shrink but do not completely eliminate large basal cell carcinomas. However, small basal cell carcinomas as in BCNS completely regress. Because these inhibitors can have severe side effects, they are often given intermittently, and thus can be given to patients for long periods of time. It may be possible to use these agents to prevent future development of BCC in patients with BCNS. Azole antifungals have been shown to inhibit this pathway as an off target effect, but small clinical trials have not revealed impressive results [84].
Curcumin
Curcumin (diferuloylmethane) is a yellow phytochemical produced by some plants, especially turmeric (Curcuma longa), a member of the ginger family (Zingiberaceae). Turmeric has been used for thousands of years in Ayurvedic and traditional Chinese medicine due to its antioxidant, anti-inflammatory, anti-tumorigenic, and antiangiogenic properties [85] but has only entered clinical trials in the last 15 years [86].
Curcumin is commonly found as an herbal supplement, an additive in cosmetics, a flavoring agent, and a food coloring. Studies show that turmeric inhibits and decreases the number of tumors in DMBA( 7,12-dimethylbenzanthracene ) induced and chronic UV initiated and 12-O-tetradecanoylphorbol-13-acetate (TPA) promoted mouse models of chemical carcinogenesis [87, 88]. While many chemotherapeutic agents may severely damage normal cells, curcumin can be safely administered in large quantities with little to no side effects, due to its preference for inducing apoptosis in highly proliferating cells [89]. As a result, curcumin could be a useful dietary additive or pharmacological treatment for the prevention of cancer incidence and mortality.
Curcumin inhibits the activity of interleukins, a group of pro-inflammatory cytokines that play vital roles in the induction of adhesion molecules, metalloproteinases, and pro-angiogenic factors. In addition, interleukins are involved in tumor invasion and angiogenesis through signaling pathways such as NF-kB, VEGF and JAK-STAT, which are inhibited by Curcumin [90–92]. The inhibition of NF-kB plays a central role in general inflammation and immune response. Suppression of COX-2 expression by inhibiting ERK activity and NF-kB activation may represent molecular mechanisms underlying this anti-inflammatory effect [93, 94]. VEGF overexpression, in particular, promotes angiogenesis and tumor growth in a number of cancers including colon, lung, and pancreas. Therefore, agents that inhibit VEGF expression have promising chemotherapeutic potential. In fact, a previous study demonstrated curcumin’s ability to inhibit VEGF expression in murine pancreatic and ovarian tumors in a similar action to Avastin (bevacizumab), an FDA-approved drug for certain cancers [95].
In 1998, Arbiser et al showed that Curcumin and its derivatives demonstrated a significant inhibition of bFGF-mediated corneal neovascularization in the mice suggesting that curcumin is a direct angiogenesis inhibitor. Furthermore they also noted effective inhibition of endothelial cell proliferation by Curcumin in a dose-dependent manner [96]. Fibroblast growth factors (FGF) are a family of growth factors involved in angiogenesis and are overexpressed in a variety of tumors.
Curcumin plays a significant role in the abrogation of a number of inflammatory and oncogenic pathways like COX-2 expression (Fig. 7.3). In a study performed in 2005, treatment of UVB-irradiated HaCaT cells with up-regulated COX-2 with curcumin strongly inhibited COX-2 mRNA and protein expression. Notably, there was also effective inhibition on UVB-induced activations of p38, MAPK, AP-1 and JNK. These results collectively suggest that curcumin may be an effective sunscreen for the protection from photo-inflammation [97]. Despite its relatively low toxicity, curcumin has poor bioavailability due to its rapid metabolism in the liver and intestinal tract [98]. As a result, until scientists can develop a method for increasing the bioavailability of curcumin, it may not be an effective chemopreventive method despite its obvious benefits.
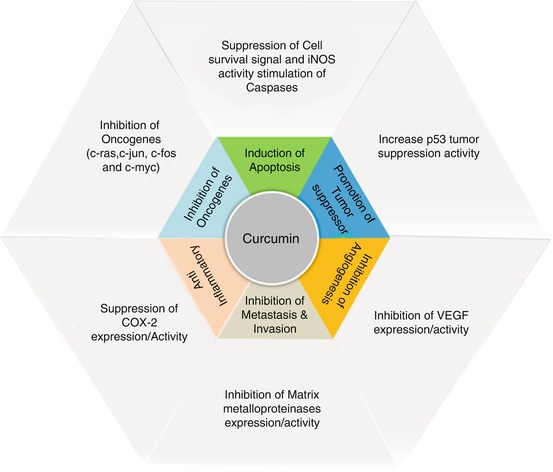
Fig. 7.3
Chemopreventive effects of Curcumin as suggested in human/ animal studies
Long-chain n-3 Polyunsaturated Fatty Acids
Long-chain n-3 polyunsaturated fatty acids (LC n-3 PUFAs), also known as Omega-3 fatty acids, are essential for normal metabolism [99]. The three types of LC n-3 PUFAs involved in human health are eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA), which have protective effects on the heart, and alpha-linoleic acid (ALA). LC n-3 PUFAs are not naturally produced in the body, and thus must be obtained through the diet. EPA and DHA are commonly found in marine oils such as fish and krill, while ALA is found in plant oils such as flaxseed, chia, and hemp. Because LC n-3 PUFAs have anti-inflammatory, antiangiogenic, and anticancer properties, their beneficial effects against cancers are widely documented [100, 101]. Numerous clinical studies support the role of n-3 PUFAs for the prevention of both melanoma and non-melanoma skin cancer (NMSC) and primarily attributes their activity to EPA and DHA [101, 102]. In addition to being an effective chemopreventive agent, studies have shown that nutritionally induced changes in tumor fatty acid composition result in increased sensitivity to chemotherapy. This effect is especially observed in chemotherapy resistant tumor lines, by causing augmented cytotoxicity to tumor cells while protecting normal cells [103].
Preclinical studies on LC n-3 PUFAs demonstrate its strong antineoplastic, apoptotic, anti-metastatic, and antiangiogenic activities due to the number of molecular factors and pathways affected. Laboratory experiments show the ability of DHA to prevent UV-induced apoptosis by increasing the expression of the anti-apoptotic Bcl-2 protein, and inhibiting the expression of human antigen R (HuR), which promotes the inflammatory properties of COX-2 [104]. The decreased expression of COX-2 also decreases B-catenin, a protein overexpressed in many cancers [105, 106]. Lastly, DHA has a pro-differentiating effect, which is integral to evade a malignant cell phenotype [105]. Other laboratory experiments demonstrate that EPA can decrease invasiveness, collagenase IV production, and metastasis in melanoma cells [107]. In addition, animal studies show that an omega-3 fatty acid rich diet significantly reduces the growth rate and angiogenesis of a human breast cancer xenograft without evidence of harmful side effects [108].
Recent clinical trials have shown the chemopreventive role of n-23 PUFAs in abrogating photo-immunosuppression [109] and decreasing inflammation [110]. Specifically, -3 PUFAs have shown antitumor activity, while -6 PUFAs and its derived eicosanoids promote anti-carcinogenesis, anti-angiogenesis, and prevent invasion [111]. Administering n-3 PUFAs decreases mucosal and epidermal response, while moderately enhancing the antitumor effect of irradiation. The magnitude of the differential effect suggests that n-3 PUFAs need to be further investigated in the clinic [112].
The mechanism underlying the antineoplastic effect of PUFAs is unclear. Initially, the idea was that n-3 PUFAs have health benefits, and do not produce dangerous levels of cytotoxic and carcinogenic oxidative products in tissues [108]. Another proposed mechanism was that lipid raft signaling proteins can be modulated in inflammation and cancer by n -3 PUFAs [113]. Recent pharmacological studies have suggested several molecular targets for the anti-inflammatory effects of omega-3 fatty acids, namely, nuclear receptor PPARc and the G protein-coupled receptor GPR120. Another proposed mechanism is that omega-3 fatty acids are converted to resolvins and protectins, lipids that have anti-inflammatory and tissue healing activity [114].
Nevertheless, further effort is needed to identify the main molecular targets of LC n-3 PUFAs in melanoma through additional well-designed human observational studies.
Vitamin A
Vitamin A and its analogs represent a group of diverse organic compounds including carotenoids and retinoids, which all have structural or biological effects similar to vitamin A [115]. In the diet, carotenoids are found in leafy green and yellow vegetables, while retinoids exist naturally in the liver and kidneys [116]. Vitamin A and its derivatives regulate a variety of essential biological processes during normal development including vision, cell proliferation and differentiation, bone tissue growth, immune function, and activation of tumor suppressor genes. While Vitamin A is a vital part of a healthy diet, its derivative, retinoids, are important in maintaining skin health and reducing susceptibility to chemical carcinogens [117] by regulating gene expression [118]. The major skin diseases that benefit from treatment with topical retinoids such as tretinoin are acne, psoriasis, disorders of keratinization, premalignant lesions such as actinic keratosis and HPV-induced tumors, and non-melanoma skin cancer [119]. Because of its apparent chemopreventive properties, patients at high risk for skin cancer due to genetic disposition (xeroderma pigmentosum), organ transplantation, or history of multiple tumors, may be treated with retinoids like etretinate or acitretin for 6 months [120].
Retinoids such as etretinate, acitretin, and isotretinoin are most effective in the treatment of precancerous lesions, such as leukoplakias, cervical dysplasia, and actinic keratosis [119]. Tazarotene, a topical retinoid, demonstrated a sustained chemopreventive effect in patients with basal cell nevus syndrome and inhibition of microscopic basal cell carcinomas even 5 months after treatment stopped [121]. These anticancer effects of applied retinoids are attributed to their impact on cellular metabolism, cell cycle progression, and inflammatory pathways. Retinoids upregulate RARβ2 [122], a tumor suppression gene, and induces the expression of p21CIP1 and p27Kip1, cell cycle regulatory proteins that cause growth arrest [123, 124]. Lastly, retinoids effectively modulate inflammatory pathways that are critical for carcinogenesis such as COX-2 [125], IL-6 and HIF-1α [126].
Despite promising in vitro and in vivo data, some prospective observational studies and randomized controlled trials on retinoids have exhibited a number of side effects. Prolonged exposure to retinoids can cause dry skin, cheilitis, conjunctivitis, and hypertriglyceridemia, effects that can exclude certain individuals from participating. Furthermore, retinoids can be teratogenic, which is a concern for women [127]. The adverse effects demonstrate the importance of finding a retinoid or treatment option that reduces these side effects, as the chemopreventive properties of retinoids are highly promising.
Vitamin D
Vitamin D is a fat-soluble vitamin that is naturally present in very few foods and is available as a dietary supplement. Most commonly, vitamin D is produced endogenously when ultraviolet rays from sunlight strike the skin and trigger vitamin D synthesis. Vitamin D plays a vital role in calcium homeostasis, cell proliferation and cell differentiation [128]. However, the vitamin D obtained from sun exposure, food, and supplements is biologically inert and must be activated inside of the body. The first hydroxylation occurs in the liver, and converts vitamin D to calcidiol (25-hydroxyvitamin D). The second occurs primarily in the kidney, and forms calcitriol (1,25-dihydroxyvitamin D) [129]. This physiologically active calcitriol is then free to bind to the vitamin D receptor (VDR) which affects gene expression by regulating transcription responses and microRNA-directed post transcriptional mechanisms [130].
1,25-dihydroxyvitamins D and analogs have been reported to inhibit the proliferation of and induce the differentiation of a wide variety of cancer cell types, including human malignant melanoma [131]. This mechanism is carried out through the VDRs, which work as tumor suppressors. Laboratory experiments show that in keratinocytes lacking VDR, there is an increased expression of oncogenes and decreased expression of tumor suppressors, which predisposes the VDR deficient mice to skin cancer formation [132]. Vitamin D3 and its analogs also demonstrate moderate antiangiogenic properties, similar to those of all-trans retinoic acid [133].
Therefore, vitamin D signaling can protect the skin from cancer formation by controlling keratinocyte proliferation and differentiation by suppressing activation of the Hedgehog signaling pathway (Hh pathway) following UVR exposure [133], facilitating DNA repair, and preventing neo-vascularization [134–136].
Analogs of vitamin D belong to the class of potent inducers of differentiation of human cancer cell lines [137]. Vitamin D and its analogs induce apoptosis when combined with retinoid acid [138] through the reduction of bcl-2 protein, a pro-apoptotic protein, and increase in p53 and p21WAF-1, both tumor suppressors [139]. Because of this synergism and known chemopreventive activity of retinoids, the combination of vitamin D analogs and retinoids could be indicated for chemoprevention of carcinomas in high risk groups (xeroderma pigmentosum, arsenical keratoses, early actinic keratoses developing in chronic radiodermitis or in patients with epidermolysis bullosa dystrophica). Lastly, when combined with various cytokines, Vitamin D has an antitumor effect [140], and may be an effective chemoprevention strategy. Despite success in animal experiments, studies with Squamous-cell carcinoma (SCC) failed to respond to 1.25(OH)2D3, a Vitamin D analog, and showed no change in VDR gene expression compared to normal keratinocytes [133]. To ameliorate this issue, perhaps some modifications of the compounds or new derivatives of vitamin D may prove more efficacious and should be introduced in clinical practice [141].
Polyphenols
Research is human cell cultures indicates extensive anticancer properties of resveratrol, a natural phenol found in red grapes, red wine, peanuts, and pines [142]. Through anti-angiogenic, pro-apoptotic, and antioxidant behavior in skin cancers, resveratrol has proved to be a successful chemopreventive agent in murine models [143–145]. Its anti-angiogenic activity is linked to the down regulation of vascular endothelial growth factor (VEGF), a critical protein in the recruitment of new blood vessels, and the up-regulation of the p53 tumor suppressor [142, 143].
Studies on human melanoma show that resveratrol also decreases expression of thrombospondin 1 (TSP1), hypoxia inducible factor-1α (HIF-1α), and toll-like receptor 4 (TLR4), proteins heavily implicated in angiogenesis [143, 146, 147]. The pro-apoptotic effects of resveratrol are linked to a down regulation of Bcl-2, an anti-apoptotic protein and NF-kB, a pro-inflammatory transcription factor [148]. The inhibition of lipoxygenase and cyclooxygenase are responsible for the antioxidant behavior of resveratrol [144], further enhancing its anticancer activity. Despite the aforementioned positive characteristics, in some situations, resveratrol has also exhibited pro-angiogenic properties such as in peri-infarct myocardium tissues [149]. Numerous studies on both animal and human cell line models have been successful in confirming the chemopreventive potential of resveratrol. However, small scale clinical trials have failed to improve the bioavailability and identify the exact mechanisms of actions of resveratrol [145]. Thus, further investigation into resveratrol and its chemopreventive properties is warranted to further understand its effects and improve its efficacy in human cancer.
Myricetin is another dietary polyphenol flavonoid that exhibits anticancer properties. Commonly found in vegetables, fruits, nuts, berries, tea, and red wine [150–152], myricetin demonstrates antioxidant and anti-angiogenic properties [153]. Similar to resveratrol, some studies show that it inhibits VEGF by directly decreasing expression of associated regulatory proteins HIF-1α, p-Akt, and ribosomal protein S6 Kinase (p70S6K) [154, 155]. Other studies point the anti-angiogenic effects of myricetin to its increased transcription of p21, a cell cycle regulatory gene that can therefore down regulate HIF-1α and VEGF expression [146, 156]. Its antioxidant behavior is related to its inhibition of COX-2 expression by targeting the NF-kB inflammatory pathway [157].
among many others including MAPK [158], Akt [159], Fyn [160], and JAK/STAT3 [161] oncogenic pathways. This widespread effect on multiple signaling pathways shows the potential of myricetin as a chemopreventive agent, and should be further investigated in human clinical trials.
A compound in the polyphenol class which has shown promise is a very low molecular weight polyphenol, Honokiol. Honokiol is the active principle of Magnolia extract [162] and is orally bioavailable compared to curcumin. It has previously demonstrated both antiangiogenic and antitumor activity in a number of preclinical studies [163]. In a study by Battle et al, honokiol was shown to cause apoptosis preferentially in cells derived from chronic lymphocytic leukemia patients sparing the normal lymphocytic cells. They noted an upregulation of Bax protein with no effect on anti-apoptotic protein bcl2 [164]. Honokiol has shown to decreases activation of NF-κB and Akt signaling mechanisms and thus a decreases in levels of NF-κB target genes like VEGF, matrix metalloproteinase-9, ICAM-1, COX-2 [165]. More recently, it has been shown to have chemopreventive activity in UV induced skin cancer. Among the established mechanisms of action of honokiol is induction of the mitochondrial enzyme sirt 3. This can act as an antioxidant in normal cells by induced mitochondrial fusion, while it can induce reactive oxygen in premalignant and malignant cells with defective mitochondria, thus potentially leading to selective death of cells with mitochondrial defects [166].
Summary
Skin cancer continues to affect an increasing number of people in the United States. Even though there is a number of commercially available natural and synthetic substance and an equally large numbers under study for chemopreventive mechanisms, the results leaves much to be desired. Further research is required to substantiate the chemopreventive effects of substances discussed in this chapter. A major obstacle to the use of chemopreventive natural products is the formulation of these compounds in a stable form that can be topically delivered to the basal layer of the epidermis and upper dermis to inhibit angiogenesis and carcinogenesis.
References
3.
Rees JR, Scot Zens M, Gui J, Celaya MO, Riddle BL, Karagas MR. Non melanoma skin cancer and subsequent cancer risk. PLoS One. 2014;9(6):e99674.PubMedPubMedCentral
4.
American Cancer Society. Cancer facts & figures 2016. Atlanta: American Cancer Society; 2016.
5.
Related posts:
 Infantile Hemangioma: New Insights on Pathogenesis and Beta Blockers Mechanisms of Action
Infantile Hemangioma: New Insights on Pathogenesis and Beta Blockers Mechanisms of Action
 Angiogenesis: General Concepts
Angiogenesis: General Concepts
 Angiogenesis and Nonmelanoma Skin Cancer
Angiogenesis and Nonmelanoma Skin Cancer
 The Role of Angiogenesis in the Development of Psoriasis
The Role of Angiogenesis in the Development of Psoriasis
 Angiogenesis and Pathogenesis of Port Wine Stain and Infantile Hemangiomas
Angiogenesis and Pathogenesis of Port Wine Stain and Infantile Hemangiomas
 Potential Role of Angiogenesis and Lymphangiogenesis in Atopic Dermatitis: Evidence from Human Studies and Lessons from an Animal Model of Human Disease
Potential Role of Angiogenesis and Lymphangiogenesis in Atopic Dermatitis: Evidence from Human Studies and Lessons from an Animal Model of Human Disease
Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


