Fig. 17.1
Domain structure for the traditional seven (I–VII) groups of vertebrate CTLs. Domain names are listed in the box
The natural killer (NK) receptor family (group V) contains a series of type II transmembrane proteins with a single CTLD expressed on NK cells, whose genes are clustered in the NK gene complex (NKC) in chromosome 12 [113]. They are collectively called killer lectin-like receptors (KLRs) and are equipped to regulate the killer activity of their expressing cells. Among them, heterodimers of CD94 and activating/inhibitory NKG2 receptors, as well as the activating NKG2D homodimer, recognize major histocompatibility complex (MHC) class I or related molecules, to distinguish target cells appropriately. The CTLDs included in these receptors have no capacity to bind carbohydrate.
Phagocytes, such as macrophages and dendritic cells (DCs), also express various kinds of CTL receptors on their cell surface for antigen capture. The mannose receptor (MR) family (group VI) contains type I transmembrane proteins with multiple CTLDs, such as the MR (CD206) and DEC-205 (CD205) [27]. They commonly consist of an N-terminal cysteine-rich domain and a fibronectin type II domain as well as eight or ten CTLDs. In contrast, the asialoglycoprotein receptor family (group II) contains type II transmembrane proteins with a single CTLD, such as DC-specific ICAM3-grabbing nonintegrin (DC-SIGN, CD209), dectin-1, dectin-2, DC immunoreceptor (DCIR), and macrophage-inducible C-type lectin (Mincle) [4, 5, 9, 33, 69]. Notably, expression of some receptors is specific and they can be markers for distinct DC subsets; langerin (CD207) on Langerhans cells (LCs) and blood DC antigen (BDCA)-2 (CD303) on plasmacytoid DCs (pDCs) [25, 99]. Most of the genes for these group II receptors are also clustered in or next to the NKC (Fig. 17.2) [55]. In contrast to KLRs, most receptors of both groups contain fully conserved CRDs binding carbohydrates, and are considered to work as pattern recognition receptors (PRRs) recognizing various pathogen-associated molecular patterns (PAMPs). Especially, dectin-1 is critical for recognition of fungal β-glucan and has a collaborative effect with Toll-like receptor (TLR) 2 on yeast-induced activation signals, whereas DC-SIGN is involved in the recognition of a variety of microorganisms including viruses, bacteria, fungi, and parasites, and suppresses the immune activation signals induced by these pathogens [13, 32, 100]. For these pathogen-recognizing CTL receptors, “CLRs” have been designated analogous to other PRRs, such as TLRs, NOD-like receptors (NLRs), and RIG-I–like receptors (RLRs) [37]. Notably, most of the CLRs are linked to the signaling cascade through the immunoreceptor tyrosine-based activating or inhibitory motif (ITAM or ITIM, respectively), which was first identified in paired KLRs [38, 102]. Hemi-ITAM in dectin-1 and ITAM of Fc receptor γ chain (FcRγ) associated with dectin-2, BDCA-2, and mincle recruit spleen tyrosine kinase (Syk) leading to activating signals, whereas ITIM in DCIR recruits Src homology two domain-containing tyrosine phosphatases (SHPs) exhibiting inhibitory effects (Fig. 17.3) [57, 74]. In addition, some of them are also able to bind endogenous self-molecules and are involved in some pathophysiological aspects. For example, DC-SIGN have a role in cellular trafficking, DC-SIGN and dectin-1 can mediate an interaction between DC and T cells, and dectin-2 is involved in ultraviolet (UV)-induced tolerance [2, 5, 33, 35]. Furthermore, dectin-1 and DCIR are involved in the development of autoimmunity in mice, and SNPs in CLEC4A and CLEC4E encoding DCIR and mincle, respectively, are reportedly associated with autoimmune arthritis [31, 47, 65, 108, 115].
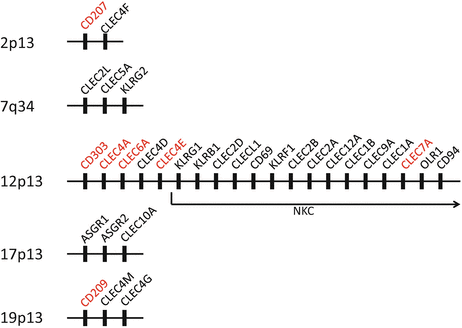
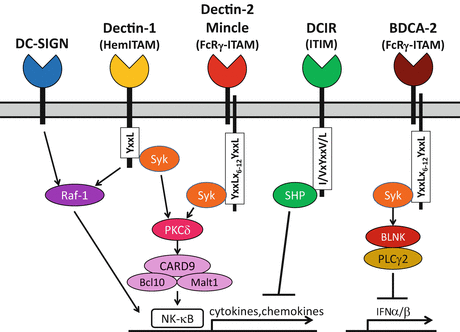
Fig. 17.2
Clustering of CLR genes. Names of the genes for CLRs discussed in this chapter are shown in red
Fig. 17.3
Representative signaling cascades through distinct intracellular motifs of CLRs (HemITAM and ITIM) or ITAM of the associating FcRγ
In this chapter, some representative CLRs are picked up and their structural features leading to the functional consequences are discussed, especially on the signaling cascades and pathogen interactions, including some impacts on cutaneous pathophysiology. Regarding the nomenclature, genes for the CLRs have formally been designated by the symbol “CLEC (C-type lectin domain containing) ”, which is approved by the Human Genome Organization Gene Nomenclature Committee (HGNC) (http://www.genenames.org/genefamilies/CLEC).
17.2 Selected CLRs
17.2.1 Langerin (CD207, CLEC4K)
Langerhans cells (LCs) are immature DCs resident in the epidermis (see Chap. 4). As sentinels, they form a honeycomb-like network and spread dendrites to the skin surface through epidermal tight junctions. Once they capture and internalize antigens, they modulate expression of adhesion molecules and chemokine receptors to migrate through dermal lymphatics into the draining lymph node and there, after maturation, they present processed antigens to the specific T cells. LCs have been defined electron-microscopically by the presence of Birbeck granules (BGs) with a “tennis-racket”-like appearance, and immunohistochemically with BG-specific Lag antibody [12, 56].
Langerin has been designated as the first surface LC marker recognized by the LC-specific monoclonal antibody (mAb) DCGM4, and subsequently isolated by the expression cloning strategy [98, 99]. Recent structural analysis has revealed that langerin stands as a stable homotrimer via the coiled-coil interaction of its neck region [28]. Langerin has no signaling motif other than the proline-rich domain for internalization in its intracellular domain. However, the presence of such an intracellular domain provides a possibility that langerin interacts with other proteins to modulate or mobilize cellular machanisms required for defense against pathogens, such as interfering viral TLR signaling [100].
As predicted by the presence of EPN (glutamate–proline–asparagine) motif in its CRD, langerin can bind mannose, fucose, and N-acetylglucosamine structures and this binding leads to internalization and transfer of the antigen into BGs, which consist of superimposed and zippered plasma membrane [92, 99]. Langerin deficiency by the W264R point mutation in humans abolished the BG formation without apparent immunodeficiency [103]. Notably, langerin expression is not specific in LCs but is also detected in a part of dermal DCs in mice [14, 39, 75]. By analysis of langerin-DTR mice, in which langerin-expressing cells were transiently depleted by diphteria toxin administration, distinct ontogeny of langerin+ dermal DCs from migrating LCs have been revealed.
Langerin recognizes human immunodeficiency virus (HIV) -1 through high-mannose structures in its envelope glycoprotein gp120 [19]. In contrast to dermal/submucosal DCs, which transmit HIV-1 to T cells through DC-SIGN, epidermal/mucosal LCs clear HIV-1 and prevent its infection to T cells through langerin without inflammation. Immunoelectron-microscopic analysis showed that HIV-1 was captured by langerin and internalized into BGs, to be finally degraded. Therefore, langerin on epidermal LCs is considered a natural barrier to HIV-1 infection. As inhibition of langerin caused HIV-1 infection in LCs and its subsequent transmission to T cells, anti-HIV therapeutics should not interfere with langerin expression or functions.
Langerin also recognizes Candida and Malassezia species through cell wall mannose structures and β-glucans [18, 96]. Mycobacterium species are also recognized and the langerin binding to their components such as lipoarabinomannan (LAM) leads to its internalization into BGs and efficient loading to CD1a for presentation to T cells [49]. Therefore, BGs might be an organelle specialized to load glycolipid antigens to CD1a.
17.2.2 DC-SIGN (CD209, CLEC4L)
Although DC-SIGN has been designated by its pivotal role on DC-T cell interaction, it is well-known as a PRR for a variety of pathogens. Indeed, it was first cloned from placenta as a HIV-1 gp120-binding protein independent of CD4 [17]. DC-SIGN stands as a tetramer and contains dileucine (LL) and YxxL internalization motif in its intracellular domain [90]. Its binding to gp120 was inhibited by mannan, D-mannose, and L-fucose and, after binding, gp120 was immediately internalized [17]. DC-SIGN does not allow HIV-1 entry into DCs, but promotes its efficient infection in trans of cells expressing CD4 and chemokine receptors [34]. DC-SIGN also functions for capturing dengue virus, a mosquito-mediated flavivirus that causes hemorrhagic fever, and is indispensable for its infection to DCs [73, 95]. Interestingly, the cytoplasmic tail of DC-SIGN is not essential for, but enhances dengue virus infection to DCs. DC-SIGN is also involved in infection of other viruses such as Ebola, cytomegaloviruses, human C-type hepatitis virus (HCV), measles, and severe acute respiratory syndrome (SARS) coronavirus, bacteria such as Helicobacter pylori and Klebsiela Pneumonae, fungi such as Candida albicans, and parasites such as Leishmania pifanoi and Schistosoma mansoni [74, 101].
DC-SIGN further works for capturing and internalizing Mycobacterium species (M. tuberculosis and BCG) through the mannose-capped mycobacterial cell wall LAM (ManLAM) [36]. In immature DCs, internalized mycobacteria or ManLAM are transported into lysosome, where they are colocalized with LAMP1. Notably, targeting DC-SIGN with soluble ManLAM inhibits mycobacteria- or lipopolysaccharide (LPS)-induced DC maturation and induces IL-10 production, suggesting that DC-SIGN signaling interferes with the TLR signaling to cause an anti-inflammatory effect. Such an effect has been shown to be mediated by a serine/threonine kinase Raf1 signaling, which does not require the cytoplasmic YxxL motif of DC-SIGN but involves formation of a distinct protein complex (LSP1, KSR, CNK, LARG, and RhoA) with DC-SIGN [42]. Raf1 signaling does not induce DC activation but modulates TLR-mediated nuclear factor (NF)-κB activation, resulting in increased and prolonged transcription of IL-10 and some other cytokines. By analysis of bronchoalveolar lavage cells, alveolar macrophages from tuberculosis patients specifically expressed DC-SIGN, and its expression was induced by M. tuberculosis independently of TLR4, IL-4, and IL-13 [97]. Indeed, accumulation of M. tuberculosis in DC-SIGN-expressing alveolar macrophages was immunohistochemically shown.
By the case-control studies, a single nucleotide polymorphism (SNP) in the promoter region of DC-SIGN, −336 C has reportedly been involved in a risk for parenteral HIV-1 infection in European Americans [68]. It has also been reported that −336 SNP is associated with severity of dengue diseases, including severe dengue fever and dengue hemorrhagic fever [86]. In tuberculosis, association of −871 SNP has further been reported by analysis of a South African population living in the areas showing a high incidence rate of tuberculosis [6]. −871G and −336A are associated with protection against tuberculosis, and their combination showed a higher frequency in non-African populations, possibly as a result of genetic adaptation to a longer history of exposure to tuberculosis.
Recently, it has been reported that the anti-inflammatory capacity of intravenous immunoglobulin (Ig) is recapitulated in mice expressing human DC-SIGN, by transfer of bone-marrow–derived macrophages or DCs treated with the Fc fragment of IgG containing glycans terminating in sialic acids (IgG-sFc) [1]. Treatment with IgG-sFc induced IL-33 production in macrophages/DCs and caused expansion of IL-4–producing basophils in their administered mice, which in turn increased expression of an inhibitory Fc receptor, FcγRIIB, on effector macrophages. Such a novel IgG-sFc–induced DC-SIGN–mediated Th2/regulatory pathway might be an endogenous mechanism for immunological homeostasis and provide a line of evidence for therapeutic effects of intravenous Ig administration on various inflammatory diseases such as Kawasaki disease, autoimmune dermatomyositis/vasculitis/neuropathy, myasthenia gravis, pemphigus vulgaris, and severe drug eruptions [88].
17.2.3 Dectin-1 (CLEC7A)
Dectin-1 was originally reported as a DC-specific lectin that had been cloned from a cDNA library of murine XS52 cells after subtraction of J774 macrophage library [5]. At first, dectin-1 was considered a costimulatory molecule because soluble recombinant dectin-1 stimulated T-cell proliferation. However, subsequently, dectin-1 was identified as a β-glucan receptor on macrophages with a different binding site from that for T cells [13]. Dectin-1 is capable of binding a major yeast cell wall component zymosan and a variety of β-glucans from fungi and plants, however, it does not recognize carbohydrates with other linkages. Although the CLEC7A gene is located within the NKC, dectin-1 expression is not observed in NK cells, but in myeloid cells including monocytes, DC, macrophages, and neutrophils [55, 114].
Dectin-1 has a cooperative role with TLR2 on zymosan-induced macrophage activation [32]. Dectin-1 and TLR2 are colocalized on the surface of zymosan-treated macrophages and the zymosan-induced activation signals require dectin-1 as well as TLR2. Notably, the YxxL hemi-ITAM (hemITAM) in the cytoplasmic tail of dectin-1 is shown to be essential for this activation. Upon zymosan-binding, this hemITAM directly recruits and activates spleen tyrosine kinase (Syk) , probably dependent on the receptor dimerization, which induces NF-κB activation through interaction with caspase recruitment domain (CARD) nine coupled with the Bcl10–Malt1 complex [46, 82]. Recently, protein kinase (PK) C-δ was identified to link Syk activation and CARD9 signaling [93]. This Syk activation also induces IL-1β secretion, through pro-IL–1β transcription and processing, mediated by the CARD9-Bcl10–Malt1 and the Malt1-ASC–caspase 8 complex, respectively [45]. The latter complex is called noncanonical caspase 8-dependent inflammasome. Additionally, zymosan-induced dectin-1–dependent signals reportedly include phospholipase (PL) Cγ 2-dependent Ca2+ influx leading to activation of mitogen-activated protein kinases (MAPKs) as well as NF-κB [109].
Furthermore, NLRP3 inflammasome is activated through dectin-1–Syk signaling in Mycobacterium abscessus (Mabc)-activated human macrophages, whereas both TLR2 and dectin-1 are required for Mabc-induced IL-1β secretion [63]. A cytoplasmic scaffold protein p62/SQSTM1 mediates this dectin 1-Syk–NLRP3 pathway, at least partly through production of reactive oxygen species (ROS).
Finally, Raf1 signaling, which regulates NF-κB activation independently of Syk, is also involved in the dectin-1 activation, as observed in case of the DC-SIGN activation [43]. Among Syk-induced cytokines, expressions of IL-1β, IL-6, IL-10, and IL-12 are enhanced but the IL-23 expression is impaired by Raf1 signaling, and, therefore, Th1 responses become dominant by dectin-1 activation. Notably, although both soluble monomer and particulate polymer of β-glucan can bind dectin-1, only the polymer can activate dectin-1 signaling by forming a so-called “phagocytic synapse,” by not only clustering the receptor but also excluding inhibitory receptors with tyrosine phosphatase activities, CD45 and CD148 [40].
Interestingly, ZAP70-deficient SKG mice reportedly developed autoimmune arthritis only in the presence of dectin-1 agonist, indicating a role of dectin-1 signaling as a trigger for manifestation of subclinical autoimmune phenotype [115]. Furthermore, zymosan-mediated dectin-1 signaling induced IL-23 production and exacerbated autoimmunity through Th1/Th17 responses in TLR2-deficient mice, whereas zymosan-mediated TLR2 signaling activated Foxp3+ regulatory T (Treg) cells through induction of IL-10 and Raldh2, an essential enzyme in retinoic acid metabolism [66]. Thus, antifungal dectin-1 signaling is considered rather aggressive and regulated by the simultaneous TLR2 signaling to prevent autoimmunity. Notably, dectn-1–deficient mice were shown to be impaired in zymosan recognition and increased in susceptibility to Candida albicans infection, whereas, in another report, they were rather more susceptible to Pneumocystis carini infection [83, 94].
In 2009, a nonsense mutation (Y238X) in the CLEC7A gene was identified in a Dutch family with chronic mucocutaneous candidiasis (OMIM#613108) [29]. The mutation was homozygous in three sisters with vulvovaginal candidiasis and/or onychomycosis and heterozygous in their parents with onychomycosis. The mutated dectin-1, which lacks C-terminal 10 amino acids, showed defective β-glucan binding and induction of cytokine productions after stimulation with β-glucan or Candida albicans. However, fungal phagocytosis and killing were shown to be unaffected and, therefore, invasive fungal infection was not observed in the patients. The allele frequency of the Y238X variant is reportedly 0.069 in the healthy Dutch population and identified carriers are all heterozygous, and the variant has never been found in the Chinese population [29]. In addition, the same variant was reported to be a risk factor of increased susceptibility to invasive aspergillosis in hematopoietic stem cell transplantation, with the highest risk in cases where the variant is present in both donors and recipients [16].
17.2.4 Dectin-2 (CLEC6A)
Dectin-2 was reported as another DC-specific lectin that had been isolated by subtraction cloning from murine XS52 cells, whereas its human homologue was originally cloned by the database search of human chromosomal sequences homologous to the exon sequences of murine DC immunoactivating receptor (DCAR) [4, 55]. DCAR was identified as the putative activating partner of DC immunoreceptor (DCIR), and the overall amino acid sequence of murine DCAR is highly homologous (61% identity) to murine dectin-2 [54]. Although the gene for dectin-2 (CLEC6A) is clustered with those for DCIR (CLEC4A) and murine DCAR (Clec4b) and located next to the NKC, dectin-2 expression is observed in various myeloid cells including monocytes, macrophages, neutrophils, and DC subsets such as LCs [55].
Indeed, murine DCAR is the first reported DC/macrophage CLR that associates with an YxxLx6–12YxxL ITAM -bearing adaptor molecule, the γ chain of FcR (FcRγ) , whereas the direct human orthologue of DCAR is absent and the functional significance of this molecule is still unknown [57]. Similar to murine DCAR, dectin-2 with a short cytoplasmic domain associates with FcRγ, a type I membrane protein with a short ectodomain that contains ITAM intracellularly and provides a docking site for Syk [85]. Dectin-2 stimulation activates the CARD 9-Bcl10–Malt1 pathway through PKCδ and induces various cytokines and chemokines mediated by NF-κB activation [80, 93]. Additionally, dectin-2 signaling activates a MAPKs pathway through PLCγ 2, as is the case with dectin-1 [41]. Most importantly, through Malt1-mediated selective c-Rel NF-κB subunit activation, dectin-2, but not dectin-1, signaling leads to Th17 responses [44].
As predicted by the presence of EPN motif in its CRD, dectin-2 binds structures with high mannose [70]. Actually, a number of pathogens including fungi such as Candida albicans, Saccharomyces cerevisiae, Aspergilus fumigatus, Cryptococcus neoformans, Paracoccidioides brasiliensis, Microsporum audouinii, and Trichophyton rubrum, as well as parasites such as Histoplasma capsulatum and Schistosoma mansoni, and Mycobacterium tuberculosis are recognized by dectin-2 [57]. Notably, the yeast and hyphal forms of Candida albicans induce different responses through dectin-2 [11, 85]. Most impressively, it has been shown by analyses of dectin-2–deficient mice that dectin-2 is essential for protective Th17 response to intravenous infection with Candida albicans [84].
By stimulation of murine bone-marrow–derived DCs with soluble schistosomal egg antigens following pretreatment with TLR ligands, NLRP3-dependent IL-1β secretion was increased through dectin-2-FcRγ-Syk signaling and ROS production [79]. Furthermore, house dust mites are also recognized by dectin-2 and induce eicosanoid production, leading to the Th2 responses [7, 8].
Interestingly, murine dectin-2 is reportedly involved in ultraviolet (UV) -induced immune regulation [2]. Although injection of soluble ectodomain of dectin-2 (sDec2) has no effect on induction of contact hypersensitivity (CHS), it inhibits UV-mediated suppression of CHS and tolerance induction. SDec2-binding CD4+ CD25+ T cells were shown to produce IL-4, IL-10, and TGFβ upon hapten stimulation and to transfer the UV-induced tolerance.
17.2.5 DCIR (CLEC4A) and DCAR (Clec4b)
DCIR was originally identified as a molecule homologous to hepatic asialoglycoprotein receptors and macrophage lectin, by selecting sequences in DC libraries following the database search of human genomic sequences using the tblastn algorithm [9]. DCIR is unique among known DC/macrophage CLRs because of the presence of I/VxxYxxV/L immunoreceptor tyrosine-based inhibitory motif (ITIM) in its cytoplasmic portion. As the ITIM-related signaling capacity of DCIR, SH 2 domain-containing tyrosine phosphatase (SHP) -1 and SHP-2 can be recruited to human DCIR upon stimulation [48, 78]. Furthermore, using murine B cells expressing a chimeric receptor containing a cytoplasmic portion of DCIR and ectodomain of FcγRIIB (DCIR-FcR), coligation of DCIR-FcR and B cell receptor (BCR) showed DCIR-ITIM-dependent inhibitory effects on BCR-mediated calcium influx and tyrosine phosphorylation of cellular proteins [52]. On the other hand, murine DCAR was identified as a molecule whose CRD shows 91% amino acid identity with that of DCIR [53]. DCAR interacts with FcRγ to stand efficiently on the cell surface and ligation of the chimeric DCAR-FcR induced activation signals into its expressing cells dependent on the ITAM in FcRγ. Thus, DCIR and DCAR are equipped as structurally and functionally paired immunoreceptors in mice, although the biological significance of their pairing has been undefined [55].
Among in vitro generated human DC subsets, a higher expression of DCIR was observed in CD14+-derived dermal-type DCs than in CD1a+-derived LC-type DCs [9]. In monocyte-derived DCs (moDCs), its expression did not change during differentiation from monocytes to immature DC, but remarkably decreased after stimulation with LPS or CD40 ligand. Notably, DCIR expression was detected not only in myeloid DCs, but also in monocytes, macrophages, B cells, neutrophils, and plasmacytoid DCs (pDCs) [55, 71]. By reverse transcription (RT)-PCR, the only reliable method to distinguish expression of DCIR and DCAR, DCAR was rather limitedly observed, strongly in spleen and lung and weakly in skin and lymph nodes, whereas DCIR was detected ubiquitously. Both increased during the bone-marrow–derived DC development, but increase of the DCAR expression preceded that of DCIR [53]. Notably, murine DC-specific mAb 33D1 recognized DCIR2, which can be a specific marker for CD8– DCs located in the red pulp and marginal zone of the spleen [23].
Despite the presence of an EPS (glutamate–proline–serine residues) motif conserved in DCIR and DCAR, which is postulated to recognize galactose residues [21], their recognizing carbohydrates have not been clarified. Notably, DCIR reportedly participates in capturing HIV-1 and its transmission to CD4+ T cells [60]. HIV-1 captured by surface DCIR is stored as intact virion and replicates de novo intracellularly in infected DCs and, subsequently, is transmitted to CD4+ T cells through induced DCIR expression on them, where HIV -1 replication is enhanced [61, 62]. These processes have been shown to be mediated by the ITIM signaling. Furthermore, DCIR can act as an attachment factor for HCV on pDCs [30]. Notably, the binding of HCV to DCIR on pDCs induced specific inhibition of TLR9-induced IFNα production, in contrast to the case of DCIR activation on DCs by anti-DCIR monoclonal antibody (mAb), which induced specific inhibition of TLR8-induced IL-12 and TNFα production [30, 71, 72].
As predicted by the presence of a tyrosine-based internalization motif overlapping with the ITIM, targeting DCIR on DCs with antigen-conjugated Abs induced clathrin-dependent receptor internalization, its trafficking into lysosomal compartments, and efficient antigen cross-presentation to CD8+ T cells [9, 59]. In contrast, targeting murine DCIR2 induced class II presentation to CD4+ T cells, including stimulation of Foxp3+ Treg cells [23, 112].
The most impressive results on in vivo significance of DCIR have been obtained by analyses of DCIR2-deficient mice. They spontaneously developed sialadenitis and enthesisis and exacerbated collagen-induced arthritis with marked expansion of DCs [31]. Furthermore, in humans, by the case-control study, a SNP in the CLEC4A gene was shown to be associated with the susceptibility to anticyclic citrulinated peptide Ab (ACPA)-negative rheumatoid arthritis (RA) [47, 65].
17.2.6 BDCA-2 (CD303, CLEC4C)
Among BDCAs, which were designated for a series of antigens recognized by mAbs raised against human CD4+ lineage– blood DCs, BDCA-2 is unique by its restricted expression on CD11c− CD123 (IL-3 receptor)bright pDCs in fresh human blood without expression on immature moDCs or CD34+-derived DCs [24]. Although murine dectin-2 was once proposed as the murine homologue of human BDCA-2, its proper murine orthologue is still undefined and murine DCAR , which lacks a human orthologue, is rather considered closely related to human BDCA-2 on structural and functional aspects [74].
In DC subpopulations, BDCA-2 expression was detected strongly in pDCs and very weakly in immature moDCs, and its expression level was downregulated after culture with IL-3 in pDCs and after maturation with LPS in moDCs [3, 25, 26]. In tonsil, BDCA-2 expression was observed on CD123+ pDCs that were primarily present in T-cell areas but not in the germinal center.
Similar to DCAR and dectin-2, BDCA-2 associates with FcRγ and cross-linking of BDCA-2 on pDCs results in intracellular Ca2+ mobilization depending on Src-family protein tyrosine kinases and on tyrosine phosphorylation of cellular proteins [15, 25]. However, unlike other FcRγ-coupled CLRs, BDCA-2 signaling does not lead to NF-κB activation through the Syk –PKCδ–CARD9 pathway, but passes through the Syk–BLNK–PLCγ 2 pathway [81]. Indeed, contrary to its predicted immunoactivating property, ligation of BDCA-2 suppressed the production of type I IFN and the secretion of TNF-related apoptosis-inducing ligand (TRAIL) in pDCs induced by various TLR agonists [25, 76]. Furthermore, inhibition of type I IFN production enhances IL-12 production in pDCs to polarize immune responses towards Th1 [26]. BDCA-2-mediated suppression of type I IFN production by CpG oligonucleotides or by influenza virus suggests that viruses target BDCA-2 for immune escape. On the other hand, the same phenomenon induced by antidouble-stranded DNA mAb plus plasmid DNA or by sera from patients of systemic lupus erythematosus (SLE) suggests that targeting BDCA-2 is beneficial for treatment of SLE.
As predicted by the presence of the triglutamate (EEE) late endosomal sorting motif in the cytoplasmic portion of BDCA-2, it was reported that targeting BDCA-2 with mAb induced internalization of the Ab complexes through phosphorylation of actin, tubulin, and clathrin, resulting in antigen presentation to CD4+ T cells [25]. However, more recently, BDCA-2 signaling has rather been shown to inhibit antigen processing and presentation to T cells by pDCs, and, furthermore, to suppress the induction of costimulatory molecules on CpG-stimulated pDCs [51].
Although BDCA-2 contains the EPN motif in its CRD similar to dectin-2, it has recently been reported that BDCA-2 recognizes asialo-galactosyl-oligosaccharides with terminal β-galactose residues and that pDCs bind to subsets of CD14+ monocytes, moDCs, and several tumor cell lines via recognition of asialo-galactosyl-oligosaccharides by BDCA-2 [77]. Accordingly, these cells can introduce the suppression signal into their binding pDCs through BDCA-2, especially on type I IFN secretion. Furthermore, BDCA-2 can bind gp120 of HIV-1 and HCV glycoprotein E2, both of which can also suppress the type I IFN secretion by pDCs [30, 67].
17.2.7 Mincle (CLEC4E)
Mincle was originally identified from murine peritoneal macrophages by subtraction cloning as a molecule specifically induced by activation of NF-IL6 transcription factor [69]. Mincle is expressed on monocytes, macrophages, neutrophils, myeloid DCs, and subsets of B cells, but not on T cells, pDCs, and NK cells [104].
Mincle associates with FcRγ dependently on the positively charged arginine residue in the transmembrane portion of mincle, as is the case with murine DCAR but is not the case with dectin-2 [110]. Mincle-FcRγ activation induces signaling through Syk –PKCδ–CARD 9–NF–κB and MAPK pathways, resulting in the induction of various cytokines and chemokines.
Related posts:

Stay updated, free articles. Join our Telegram channel

Full access? Get Clinical Tree


